
Abstract
Perea, Duñabeitia, and Carreiras (Journal of Experimental Psychology: Human Perception and Performance 34:237–241, 2008) found that LEET stimuli, formed by a mixture of digits and letters (e.g., T4BL3 instead of TABLE), produced priming effects similar to those for regular words. This finding led them to conclude that LEET stimuli automatically activate lexical information. In the present study, we examined whether semantic activation occurs for LEET stimuli by using an electrophysiological measure called the N400 effect. The N400 effect, also known as the mismatch negativity, reflects detection of a mismatch between a word and the current semantic context. This N400 effect could occur only if the LEET stimulus had been identified and processed semantically. Participants determined whether a stimulus (word or LEET) was related to a given category (e.g., APPLE or 4PPL3 belongs to the category “fruit,” but TABLE or T4BL3 does not). We found that LEET stimuli produced an N400 effect similar in magnitude to that for regular uppercase words, suggesting that LEET stimuli can access meaning in a manner similar to words presented in consistent uppercase letters.
Similar content being viewed by others
Humans possess the incredible ability to identify words, irrespective of font type, size, or relative letter position (for examples, see, e.g., Dehaene, Cohen, Sigman, & Vinckier, 2005; Velan & Frost, 2007). Most models of visual word recognition attempt to explain these (and other) phenomena by considering multiple levels of activation, including orthographic, lexical, phonological, and semantic (e.g., Allen, Smith, Lien, Kaut, & Canfield, 2009; Balota, Yap, & Cortese, 2006; Coltheart, Rastle, Perry, Langdon, & Ziegler, 2001; McClelland & Rumelhart 1981). These models typically assume a continuous flow of information from recognizing orthographic form to retrieving meaning, in either a cascaded or a parallel processing mode. Despite diverse assumptions proposed in variations of those models, they generally agree that visual word recognition is mediated by the perceptibility of words (see Lupker, 2005, for a review).
A study by Perea , Duñabeitia, and Carreiras (2008) demonstrated that visual word recognition is not disrupted by displaying words with characters that share similar configural features with the letters that constitute the words. Perea et al. used LEET stimuli, in which certain letters are replaced by letter-like digits (e.g., the digit “3” for the letter “E,” or R34D1NG instead of READING). They argued that if detecting the shapes of individual letters is sufficient to produce lexical activation, then letter-like digits or letter-like symbols should produce lexical activation just like regular words. To test this claim, Perea et al. used a masked-priming paradigm. They presented a forward mask (a row of #s) for 500 ms, followed by a 50-ms prime in the center location. Immediately after the offset of the prime, the target appeared in the same location and remained on screen until participants made a response. To reduce the spatial overlap between prime and target, Perea et al. presented the primes and targets in different font sizes (10-point Courier vs. 12-point Courier, respectively). Participants were to determine whether the target was a word (a lexical-decision task). In Experiment 1, the prime was either the same target word (e.g., MATERIAL for the target MATERIAL), the corresponding LEET for that target (e.g., M4T3R14L), or stimuli with letters replaced with corresponding symbols (letter-like symbols; e.g., M∆T€R!∆L) or control letters (replacing the original letters with other letters, randomly selected; e.g., MOTURUOL for the target MATERIAL).
Perea et al. (2008) found that response times (RTs) to the target word were significantly faster when the word was primed by LEETs and symbols than when it was primed by the control letters (i.e., a priming effect). Most importantly, both LEETs and symbols produced mean RTs similar to those for the target word prime. They further found, in Experiment 2, that both LEETs and symbols produced faster RTs than their corresponding control conditions, in which other non-letter-like numbers and symbols, respectively, were used (e.g., M6T2R76L for the target MATERIAL in the control LEET condition; MT%R?L in the control symbol condition). They concluded that “it is visual similarity rather than the status of the leet digits as numbers that seems to be responsible for the leet priming effect” (p. 239).
Perea et al.’s (2008) findings with the masked-priming paradigm seem to support the conclusion that, in the absence of a top-down context, letter-like digits (and symbols) embedded in a word are sufficient to trigger lexical activation. Nevertheless, some studies have suggested that priming effects result from prelexical processing (i.e., affecting stages prior to lexical activation; see Hutchison, Neely, Neill, & Walker, 2004; Masson & Isaak, 1999). In other words, the similarity of the visual shapes of LEET stimuli to actual letters may have affected only orthographic, but not lexical, processing of the target word. Evidence favoring the sublexical account mainly comes from the finding of similar repetition priming effects for words and nonwords in the lexical decision task, even though nonwords should not benefit from lexical processing of primes (e.g., Masson & Isaak, 1999). Thus, the advantage for LEET stimuli over the nonword control primes (e.g., M4T3R14L vs. MOTURUOL for the target word MATERIAL) in Perea et al. could arise merely from less visual impairment in the LEET condition than in the other stimulus conditions (e.g., Davis & Lupker, 2006; Horemans & Schiller, 2004; see Forster, 1998, for a review).
Because the priming effect with LEET stimuli observed in Perea et al. (2008) cannot be unambiguously attributed to lexical activation, this highlights a need for converging evidence. Because it is unclear whether priming results from lexical activation, we instead examined the semantic activation of LEET stimuli using a category judgment task—determining whether a single stimulus (either a word or a LEET stimulus) was related or unrelated to the category provided prior to that block. It has been suggested that semantic-relatedness judgments, regardless of whether they are related or unrelated, are a result of lexical activation (e.g., Besner, Smith, & MacLeod, 1990). Accordingly, if we were to find any evidence of semantic activation, we could infer the occurrence of lexical activation.
In addition to examining semantic activation using behavioral data (e.g., RTs), we used event-related potential (ERP) measures. ERPs provide online, continuous measures of meaning extraction and often reveal evidence of deeper processing than is apparent in behavioral data. For instance, Heil, Rolke, and Pecchinenda (2004) found a modulation of ERP amplitudes by the semantic relatedness of prime and probe words, even when there was no semantic priming effect in RTs (e.g., Rolke, Heil, Streb, & Hennighausen, 2001; see also Vogel, Luck, & Shapiro, 1998, for an example of ERPs elicited by semantic activation even when participants could not report the targets in the attentional blink task). By examining the ERP components associated with semantic activation, it is possible to determine whether LEET stimuli trigger semantic activation just like regular words do.
We used the N400 component, a negative-going brain potential that occurs around 400 ms after the onset of potentially meaningful stimuli (e.g., words). This component is also called the mismatch negativity, because it is known to be elicited when a person notices a stimulus that is incongruent with the current semantic context (see Kutas & Van Petten, 1988, for a review). For instance, after one sees the word SPORTS, the word APPLE (unrelated) would elicit a much larger negative ERP than would the word TENNIS (related) 400–600 ms after word onset. Therefore, the N400 effect can be quantified as the average difference in brain potentials between words that are related and unrelated to the current semantic context (N400 effect = unrelated word ERPs – related word ERPs).
The critical point is that the N400 effect can be used as an indicator that a person has extracted a word’s meaning. That is, the N400 effect occurs only when participants detect a semantic mismatch, indicating processing even deeper than lexical activation. It has been suggested, in fact, that the N400 effect is a more sensitive measure of semantic activation than are behavioral measures (e.g., Heil et al., 2004; Rolke et al., 2001; Vogel et al., 1998). As a concrete example, Lien, Ruthruff, Cornett, Goodin, and Allen (2008) used N400 effects to determine whether people can extract word meaning while central attention is devoted to another task. Participants performed a tone–pitch Task 1 and a semantic-relatedness Task 2 (i.e., whether the Task 2 word was related to a previously presented context word/category). The critical finding was that the N400 effect declined sharply under dual-task conditions. They concluded that semantic activation of visual words is impaired while central attention is allocated to another task.
The present study
In the present study, we used ERPs (i.e., the N400 effect) to assess whether semantic activation occurs for LEET stimuli just as strongly as it does for regular words. Thus, regular words were included to provide a baseline for semantic activation. We adopted Lien et al.’s (2008) category judgment task, in which participants memorized a category name prior to each block (e.g., fruit) and then determined whether a series of single stimuli (e.g., APPLE for the regular word or 4PPL3 for the LEET stimulus) were related or unrelated to that category. Stimulus type (word vs. LEET) varied within blocks.
Our main interest was the semantic activation of words and LEET stimuli, as indicated by the N400 effect (the difference in ERPs between unrelated and related words). The semantic relatedness effect (the difference between unrelated and related words) on RTs does not provide a clear picture regarding semantic activation for words and LEET stimuli, since related and unrelated responses are made with different response fingers. Thus, the effect on RTs could reflect a modulation of response decision processes (see the Discussion regarding the problems with interpreting semantic activation using behavioral data). Furthermore, since we compared semantic activation between words and their corresponding LEET stimuli, word frequency and word length should have little influence on the comparison between them.
We expected to observe a large N400 effect for regular words, as had previously been shown in single-task conditions (e.g., Lien et al., 2008). The main question for the present study was whether similar N400 effects would also be observed for LEET stimuli formed by letters and digits. If LEET stimuli are processed like real words (i.e., if they can access word meaning), then LEET stimuli should produce N400 effects similar to those from regular words. Such a result not only would imply semantic activation for the LEET stimuli, but also would provide converging evidence for Perea et al.’s (2008) claim of full lexical activation for LEET stimuli.
Method
Participants
A group of 24 undergraduates (native English speakers) at Oregon State University participated in this experiment. The data from two of these participants were excluded due to excessive eye movement artifacts in the electroencephalographic (EEG) data (see below). The remaining 22 participants (16 females, six males) had a mean age of 20 years (range: 18–27).
Apparatus, stimuli, and procedure
Stimuli were displayed on a 19-in. monitor and were viewed from a distance of about 55 cm. The category presented prior to each block was in lowercase, whereas the stimulus for each trial was a string of uppercase letters (words) or a mixture of letters and digits (LEET stimuli) printed in white, against a black background, in the center of the screen. LEET stimuli were formed by changing some letters in a word using digits that were similar in shape to their corresponding uppercase letters. That is, the digit 1 was used for the letter L, the digit 3 for the letter E, the digit 4 for the letter A, the digit 5 for the letter S, the digit 6 for the letter G, and the digit 0 for the letter O. The digit substitutions ranged from 30% to 64% (mean: 43%) across letter positions. Each letter and digit subtended a visual angle of approximately 1.15º in width × 1.25º in height.
Each trial started with a fixation cross in the screen center for 1,200 ms, which was then replaced by the stimulus until a response was made. Next, auditory feedback (a tone on error trials, silence on correct trials) was presented for 100 ms. The next trial then began with the fixation display. The participant’s task was to indicate whether the stimulus was related or unrelated to the category for that block by pressing the leftmost response-box button for related and the rightmost button for unrelated stimuli. They were also told that some words would be formed by digits and letters, which they should treat like regular words and determine the semantic relatedness to the current category. Speed and accuracy were emphasized equally.
A total of 20 categories were used for the experimental blocks, and two categories were used for the practice blocks, taken from Lien et al. (2008; see the Appendix for the complete list).Footnote 1 Each participant performed two sessions. The first session contained one practice block of 36 trials, randomly selected from the two practice-block categories, followed by 20 experimental blocks of 36 trials each (nine related words, nine related LEETs, nine unrelated words, and nine unrelated LEETs, randomly determined). The second session consisted of the same 20 categories of experimental blocks as the first session, except that the related/unrelated words and LEET stimuli were different. The order of categories for the experimental blocks was randomly determined for each participant. For each participant, each word and LEET stimulus appeared twice throughout the whole experiment—once for the related list and once for the unrelated list, in a random order. Participants completed these two sessions within a single visit to the lab and were given breaks between blocks and sessions.
EEG recording and analyses
Electroencephalographic (EEG) activity was recorded from electrodes F3, Fz, F4, C3, Cz, C4, P3, Pz, P4, O1, and O2. These sites and the right mastoid were recorded in relation to a reference electrode at the left mastoid. The EEGs were then re-referenced offline to the average of the left and right mastoids. A horizontal electrooculogram (HEOG) was recorded bipolarly from electrodes at the outer canthi of both eyes, and a vertical electrooculogram (VEOG) was recorded from electrodes above and below the midpoint of the left eye. The electrode impedance was kept below 5 kΩ. EEG, HEOG, and VEOG were amplified using Synamps2 (Neuroscan) with a gain of 2,000 and a bandpass of 0.1–70 Hz, and were digitized at 250 Hz.
Trials with possible ocular and movement artifacts were identified using a threshold of ±75 μV for a 1,400-ms epoch beginning 200 ms before stimulus onset to 1,200 ms after stimulus onset. Each of these artifact trials was then inspected manually. This procedure led to the rejection of 7% of the trials, with no more than 25% rejected for any individual.
Results
In addition to trials with ocular artifacts, trials were excluded from analyses of the behavioral data (RTs and proportions of errors) and the ERP data if the RT was less than 100 ms or greater than 2,000 ms (0.4% of trials exceeded these cutoff values). Incorrect-response trials were also excluded from the RT and ERP analyses. Analyses of variance (ANOVA) were used for all statistical analyses. Whenever appropriate, p values were adjusted using the Greenhouse–Geisser epsilon correction for nonsphericity.
Behavioral data analyses
The ANOVAs on RTs and proportions of errors were conducted as a function of stimulus type (word vs. LEET) and semantic relatedness (related vs. unrelated). Table 1 shows the mean RTs and proportions of errors for each condition. The analyses revealed that the overall RT was 41 ms longer for LEET than for word stimuli, F(1, 21) = 103.76, p < .0001, η p 2 = .83, and the mean RT was 44 ms longer for unrelated stimuli (677 ms) than for related stimuli (633 ms), F(1, 21) = 44.62, p < .0001, η p 2 = .68. The semantic-relatedness effect (unrelated – related) on RTs was larger for LEET stimuli than for words (58 vs. 29 ms, respectively), F(1, 21) = 22.91, p < .0001, η p 2 = .52. Further t test analyses revealed that the semantic-relatedness effect was significant for both LEET stimuli, t(21) = 7.31, p < .0001, and words, t(21) = 4.58, p < .001.
For the proportion-of-error data, LEET stimuli produced higher error rates than did word stimuli (.085 vs. .072), F(1, 21) = 27.73, p < .0001, η p 2 = .57. The error rate was also higher for related (.100) than for unrelated (.056) stimuli, F(1, 21) = 28.83, p < .0001, η p 2 = .58. However, the semantic-relatedness effects on error rates were similar for LEET stimuli and for words (−.047 vs. –.041, respectively), F < 1.0. As in the RT data, further t test analyses revealed that the semantic-relatedness effects were significant for both LEET stimuli, t(21) = 5.08, p < .0001, and words, t(21) = 4.97, p < .0001.
ERP analyses
The averaged ERP waveforms were time-locked to stimulus onset. For each stimulus type (word vs. LEET), difference waves were constructed by subtracting the ERP waveforms elicited by stimuli related to the category from the ERP waveforms elicited by stimuli unrelated to the category (i.e., the N400 effect). We collapsed across the three frontal electrode sites (F3, Fz, and F4), the three central electrode sites (C3, Cz, and C4), and the three parietal electrode sites (P3, Pz, and P4). Following Lien et al. (2008; see also Vogel et al., 1998), the mean amplitude of the N400 effect was measured from 400 to 600 ms after stimulus onset, relative to the 200-ms baseline period before stimulus onset. This is the time window during which the N400 effect is typically maximal (see Fig. 1). An ANOVA on the N400 amplitudes (difference waveforms = unrelated ERPs – related ERPs) was conducted as a function of stimulus type (word vs. LEET) and electrode site (frontal [F3, Fz, F4] vs. central [C3, Cz, C4] vs. parietal [P3, Pz, P4]). Figure 2 shows the grand average waveforms for related and unrelated stimuli for these electrodes, and Fig. 3 shows the mean N400 amplitudes.

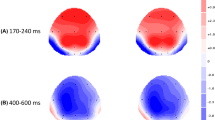
Scalp topographies of the N400 effect (difference = unrelated ERPs – related ERPs) for word and LEET stimuli during the time window 400–600 ms after stimulus onset. ERP: event-related potential

Grand average event-related brain potentials as a function of semantically related and unrelated word and LEET stimuli at frontal electrode sites (data collapsed across F3, Fz, and F4), central electrode sites (data collapsed across C3, Cz, and C4), and parietal electrode sites (data collapsed across P3, Pz, and P4). Negative is plotted upward, and time zero represents stimulus onset. The baseline period was 200 ms prior to stimulus onset.

Grand average differences in event-related brain potentials, formed by subtracting semantically related trials from semantically unrelated trials (i.e., the N400 effect), for word and LEET stimuli at frontal electrode sites (data collapsed across F3, Fz, and F4), central electrode sites (data collapsed across C3, Cz, and C4), and parietal electrode sites (data collapsed across P3, Pz, and P4). The unfilled rectangular boxes indicate the time window used to assess the N400 effect (400–600 ms after stimulus onset). Negative is plotted upward, and time zero represents stimulus onset. The baseline period was 200 ms prior to stimulus onset.
The N400 effects were similar for both words and LEET stimuli (−3.568 vs. –3.848 μV, respectively), F(1, 21) = 1.51, p = .2329, η p 2 = .03. Further t test analyses revealed that the N400 effect was significantly greater than zero for both words, t(21) = −12.88, p < .0001, and LEET stimuli, t(21) = −13.35, p < .0001. Although the overall N400 effect was much larger for the central sites (−4.050 μV) than for the frontal and parietal sites (−3.585 vs. –3.488 μV), F(2, 42) = 7.23, p < .01, η p 2 = .26, the difference in the N400 effects between words and LEET stimuli did not interact with electrode site, F < 1.0.
Discussion
In the present study, we used the N400 effect to examine whether semantic activation occurs for LEET stimuli (formed by letters and digits) just as strongly as for regular words. In each trial, participants made a category judgment on either the LEET stimuli or regular words (formed by uppercase letters), intermixed within blocks. An advantage of using a single-stimulus presentation in our study is that it eliminated unwanted effects of extraneous stimuli, such as the possible forward-masking interference produced by the priming paradigm (e.g., Forster, 1998). The critical finding was that the N400 effects at all three electrode sites (frontal, central, and parietal) were similar in magnitude for both LEET and regular words, suggesting that semantic activation occurs for LEET stimuli just as strongly as for regular words. Since it has been suggested that semantic activation is a result of lexical activation (e.g., Besner et al., 1990), we concluded that, in the presence of a top-down context (e.g., semantic categories), letter-like digits embedded in a word are able to activate both lexical and semantic information.
Another interesting finding of the present study was the apparent trend toward earlier N400 onsets for word than for LEET stimuli (see Fig. 3). Thus, even though the digits in LEET stimuli can be encoded in a letter-like manner and subsequently may activate semantic information, the regular words have faster access to the mental lexicon due to greater familiarity. The behavioral data (e.g., RTs) are also consistent with this claim, showing that faster semantic-relatedness judgments occur for words than for LEET stimuli. This finding seems to suggest that processing stages prior to semantic activation (e.g., encoding) might be delayed for LEET stimuli, but that semantic analysis of the encoded letter representations is not affected at all.
We noted earlier that the use of behavioral data (e.g., RTs) opens the door to numerous alternative explanations for the priming effect with LEET stimuli, such as prelexical processing or less-disruptive visual similarity between prime and target, rather than the lexical-activation interpretation favored by Perea at al. (2008). Likewise, our behavioral data may not provide clear evidence for semantic activation in LEET stimuli. We found a positive semantic-relatedness effect on RTs (slower for unrelated than for related stimuli), but a negative effect on errors (smaller error rates for unrelated than for related stimuli), indicating a speed–accuracy trade-off. This finding underscores another major limitation of behavioral measures—they are sensitive to response bias (e.g., Green & Swets, 1966). On the positive side, however, it also highlights one major advantage of N400 measures, as we used in the present study; that is, they more specifically reflect the buildup of semantic activation, with little constraint from decision making (e.g., Heil et al., 2004; Rolke et al., 2001; for a review, see Kutas & Federmeier, 2011).
Our N400 data provided clear evidence for semantic activation in LEET stimuli. Alternatively, the activation could be due to top-down expectancy, since participants already knew what category to look for in each block. Although we always presented a single stimulus (either a LEET stimulus or a regular uppercase word) in each trial, those stimuli were assessed in terms of their fit within a semantic category presented prior to each block. In this light, the finding that LEET stimuli elicited an N400 effect similar in magnitude to that for regular words suggests that the N400 effect is not an indicator of the perceptual integration of lexicality (because the mixture of digits and letters in LEET stimuli would have disrupted the encoding of lexical representations, with little semantic information being activated). Rather, it more likely reflects a postlexical process that is driven by integration of a context, such as categories (e.g., Brown & Hagoort, 1993; Kutas & Federmeier, 2011; but see Deacon, Hewitt, Yang, & Nagata, 2000, for a case in which the N400 effect occurred without conscious awareness of word identity).
One constraint of our study is that the effects of letter versus digit position in LEET stimuli and the number of digit replacements in the LEET stimuli may have played roles in semantic activation.Footnote 2 For instance, as was observed by Jordan, Thomas, Patching, and Scott-Brown (2003), when words were presented in passages of text, visually degrading the exterior letters of a word (the first and last positions, such as “d_ _k” for the word “dark”) slowed reading rates more than did visually degrading interior letters (all letter positions that lay between the first and last positions, such as “_ar_” for the word “dark”). They therefore argued that the exterior letters of words play a major role in visual word recognition (see also Jordan, Thomas, & Scott-Brown, 1999, with presentation of single words rather than passages). In the present study, about 55% of the LEET stimuli replaced at least one of the exterior letters with a digit (30% and 34%, respectively, of the first and last letters were replaced, and both positions were replaced for 9% of stimuli). If semantic access was restricted to digit replacements only in interior locations or only in exterior locations, one would expect the N400 effect elicited by the LEET stimuli to be smaller than the effect elicited by regular words. In contrast to this prediction, our results showed that the N400 effect for LEET stimuli was about 8% larger in magnitude than the effect for regular words. Thus, the present N400 effect for LEET stimuli was not due solely to an “exterior-letter effect.” The present LEET results, along with Jordan and colleagues’ earlier letter-position work and the case-mixing results of Allen et al. (2009) and Lien, Allen, and Crawford (2012), suggest that “coarse-scale” information in words (i.e., the cursory shape of the whole word; Jordan et al., 2003) is involved in visual word recognition in addition to individual letter identities. This appears to be the case, because all of these studies suggest that the physical identity of individual letters is not the only information used to encode words.
In sum, we have demonstrated that LEET stimuli gain lexical access and activate semantic information using electrophysiological measures (i.e., N400 effects). It is clear that the exact identity of the components in a word, such as digits instead of letters, does not hinder semantic activation, even though the encoding is slower for LEET than for regular words. Thus, we extended Perea et al.’s (2008) priming results to a category task and showed that the processing equivalence applies even to deeper levels of word processing (i.e., access to word meaning).
Notes
In order to form LEET stimuli, it was necessary to replace some words from Lien et al.’s (2008) list. It should be noted that it was very difficult to generate items for each category that could be substituted with digits. Therefore, instead of requiring LEET stimuli to have at least three digits, as in Perea et al.’s (2008) study, we required at least one digit.
We did not design our study to examine the effect of the number or position of digit replacements in LEET stimuli. Across all of the LEET stimuli, 14%, 50%, 30%, and 6% contained one, two, three, and four digits, respectively (additionally, one LEET stimulus had five digits, and one had six digits). The percentages of digit replacement in Positions 1–9 of the LEET stimuli ranged from 30% to 64%. Since the design was unbalanced, analyzing the N400 effects and the behavioral data as a function of these two variables would be biased. Most importantly, breaking down N400 effects as a function of these two variables would result in small samples, leaving noisy data.
References
Allen, P. A., Smith, A. F., Lien, M.–. C., Kaut, K. P., & Canfield, A. (2009). A multistream model of visual word recognition. Attention, Perception, & Psychophysics, 71, 281–296. doi:10.3758/APP.71.1.281
Balota, D. A., Yap, M. J., & Cortese, M. J. (2006). Visual word recognition: The journal from features to meaning (a travel update). In M. J. Traxler & M. A. Gernsbacher (Eds.), Handbook of psycholinguistics (2nd ed., pp. 285–375). Amsterdam, The Netherlands: Academic Press.
Besner, D., Smith, M. C., & MacLeod, C. M. (1990). Visual word recognition: A dissociation of lexical and semantic processing. Journal of Experimental Psychology. Learning, Memory, and Cognition, 16, 862–869. doi:10.1037/0278-7393.16.5.862
Brown, C., & Hagoort, P. (1993). The processing nature of the N400: Evidence from masked priming. Journal of Cognitive Neuroscience, 5, 34–44.
Coltheart, M., Rastle, K., Perry, C., Langdon, R., & Ziegler, J. C. (2001). DRC: A dual route cascaded model of visual word recognition and reading aloud. Psychological Review, 108, 204–256. doi:10.1037/0033-295X.108.1.204
Davis, C. J., & Lupker, S. J. (2006). Masked inhibitory priming in English: Evidence for lexical inhibition. Journal of Experimental Psychology. Human Perception and Performance, 32, 668–687. doi:10.1037/0096-1523.32.3.668
Deacon, D., Hewitt, S., Yang, C.-M., & Nagata, M. (2000). Event-related potential indices of semantic priming using masked and unmasked words: Evidence that the N400 does not reflect a post-lexical process. Cognitive Brain Research, 9, 137–146.
Dehaene, S., Cohen, L., Sigman, M., & Vinckier, F. (2005). The neural code for written words: A proposal. Trends in Cognitive Sciences, 9, 335–341.
Forster, K. I. (1998). The pros and cons of masked priming. Journal of Psycholinguistic Research, 27, 203–233. doi:10.1023/A:1023202116609
Green, D. M., & Swets, J. A. (1966). Signal detection theory and psychophysics. New York, NY: Wiley.
Heil, M., Rolke, B., & Pecchinenda, A. (2004). Automatic semantic activation is not myth: Semantic context effects on the N400 in the letter-search task in the absence of response time effects. Psychological Science, 15, 852–857. doi:10.1111/j.0956-7976.2004.00766.x
Horemans, I., & Schiller, N. O. (2004). Form-priming effects in nonword naming. Brain and Language, 90, 465–469. doi:10.1016/S0093-934X(03)00457-7
Hutchison, K. A., Neely, J. H., Neill, W. T., & Walker, P. B. (2004). Is unconscious identity priming lexical or sublexical? Consciousness and Cognition, 13, 512–538.
Jordan, T. R., Thomas, S. M., Patching, G. R., & Scott-Brown, K. C. (2003). Assessing the importance of letter pairs in initial, exterior, and interior positions in reading. Journal of Experimental Psychology. Learning, Memory, and Cognition, 29, 883–893. doi:10.1037/0278-7393.29.5.883
Jordan, T. R., Thomas, S. M., & Scott-Brown, K. C. (1999). The illusory-letters phenomenon: An illustration of graphemic restoration in visual word recognition. Perception, 28, 1413–1416.
Kutas, M., & Federmeier, K. D. (2011). Thirty years and counting: Finding meaning in the N400 component of the event-related brain potential (ERP). Annual Review of Psychology, 62, 621–647. doi:10.1146/annurev.psych.093008.131123
Kutas, M., & Van Petten, C. (1988). Event-related brain potential studies of language. In P. K. Ackles, J. R. Jennings, & M. G. H. Coles (Eds.), Advances in psychophysiology (Vol. 3, pp. 139–187). Greenwich, CT: JAI Press.
Lien, M.–. C., Allen, P. A., & Crawford, C. (2012). Electrophysiological evidence of different loci for case mixing and word frequency effects in visual word recognition. Psychonomic Bulletin & Review, 19, 677–684. doi:10.3758/s13423-012-0251-9
Lien, M.–. C., Ruthruff, E., Cornett, L., Goodin, Z., & Allen, P. A. (2008). On the non-automaticity of visual word processing: Electrophysiological evidence that word processing requires central attention. Journal of Experimental Psychology. Human Perception and Performance, 34, 751–773. doi:10.1037/0096-1523.34.3.751
Lupker, S. J. (2005). Visual word recognition: Theories and findings. In M. J. Snowling & C. Hulme (Eds.), The science of reading: A handbook (pp. 39–60). Oxford, UK: Blackwell.
Masson, M. E. J., & Isaak, M. I. (1999). Masked priming of words and nonwords in a naming task: Further evidence for a nonlexical basis for priming. Memory & Cognition, 27, 399–412. doi:10.3758/BF03211536
McClelland, J. L., & Rumelhart, D. E. (1981). An interactive activation model of context effects in letter perception: I. An account of basic findings. Psychological Review, 88, 375–407. doi:10.1037/0033-295X.88.5.375
Perea, M., Duñabeitia, J. A., & Carreiras, M. (2008). R34D1NG W0RD5 W1TH NUMB3R5. Journal of Experimental Psychology. Human Perception and Performance, 34, 237–241. doi:10.1037/0096-1523.34.1.237
Rolke, B., Heil, M., Streb, J., & Hennighausen, E. (2001). Missed prime words within the attentional blink evoke an N400 semantic priming effect. Psychophysiology, 38, 165–174.
Velan, H., & Frost, R. (2007). Cambridge University versus Hebrew University: The impact of letter transposition on reading English and Hebrew. Psychonomic Bulletin & Review, 14, 913–918. doi:10.3758/BF03194121
Vogel, E. K., Luck, S. J., & Shapiro, K. L. (1998). Electrophysiological evidence for a postperceptual locus of suppression during the attentional blink. Journal of Experimental Psychology. Human Perception and Performance, 24, 1656–1674. doi:10.1037/0096-1523.24.6.1656
Acknowledgments
We thank Cathleen Moore, Eric Ruthruff, and two anonymous reviewers for comments on earlier versions of the manuscript. We also thank Andrew Morgan for providing technical support.
Author information
Authors and Affiliations
Corresponding author
Appendix: Category labels (in bold) and their members in words and LEET stimuli used in the experiment
Appendix: Category labels (in bold) and their members in words and LEET stimuli used in the experiment
The unrelated words were selected from different categories, with the restriction that each word and LEET stimulus appeared exactly once in the related condition and once in the unrelated condition. The last two categories (emotion/expression/feeling and sports) were used for practice blocks. However, each participant received only one of these two categories, randomly determined.
Mammals | Body Parts | Transportation | |||
Word | LEET | Word | LEET | Word | LEET |
GORILLA | G0R1LL4 | EAR | 34R | TRAWLER | TR4WL3R |
PIG | P16 | TONGUE | T0N6U3 | CART | C4RT |
DEER | D33R | SHIN | 5H1N | KAYAK | K4Y4K |
ZEBRA | Z3BR4 | HEEL | H33L | PLANE | PL4N3 |
RABBITT | R4881TT | FOOT | F00T | YACHT | Y4CHT |
GIRAFFE | 6IR4FF3 | LEG | L3G | CAR | C4R |
SHEEP | SH33P | EYE | 3Y3 | TRAILER | TR41L3R |
WOLF | W0LF | NECK | N3CK | SHIP | SH1P |
TIGER | T163R | HEAD | H34D | WAGON | W460N |
APE | 4P3 | ELBOW | 3LB0W | TRAM | TR4M |
BEAVER | B34V3R | WRIST | WR15T | SLED | 5L3D |
COYOTE | C0Y0T3 | CHEEK | CH33K | CANOE | C4N03 |
BEAR | 834R | KNEE | KN33 | VAN | V4N |
CHICKEN | CH1CK3N | TOE | T03 | TRAIN | TR41N |
GOAT | GO4T | ANKLE | 4NKL3 | BIKE | B1K3 |
CAT | C4T | ARM | 4RM | BOAT | B04T |
MOOSE | M00S3 | JAW | J4W | BUS | 8U5 |
COW | C0W | FINGER | F1N63R | TROLLEY | TR0LL3Y |
Birds | Bugs | Clothing | |||
Word | LEET | Word | LEET | Word | LEET |
HAWK | H4WK | GNAT | 6N4T | SHORTS | 5H0RT5 |
PIGEON | P1630N | SLUG | 5LU6 | CAP | C4P |
FALCON | F4LC0N | BEE | B33 | SHOE | 5H03 |
HERON | H3R0N | WASP | W45P | BOOT | 800T |
SPARROW | 5P4RR0W | LICE | L1C3 | ROBE | R083 |
ROBIN | R0B1N | ANT | 4NT | HAT | H4T |
RAVEN | R4V3N | CRICKET | CR1CK3T | GLOVE | 6L0V3 |
SEAGULL | S34GULL | TERMITE | T3RM1T3 | JACKET | J4CK3T |
CRANE | CR4N3 | BEETLE | 833T13 | SKIRT | 5K1RT |
SWAN | 5W4N | HORNET | H0RN3T | JEANS | J34N5 |
PERROT | P3RR0T | CICADA | C1C4D4 | SOCK | 50CK |
PARROT | P4RR0T | APHID | 4PH1D | SWEATER | 5W34T3R |
CHICKEN | CH1CK3N | FLEA | FL34 | HOSE | H053 |
PELICAN | P3L1C4N | SPIDER | 5P1D3R | SHIRT | 5H1RT |
PEACOCK | P34C0CK | FIREFLY | F1R3FLY | DRESS | DR355 |
GOOSE | G00S3 | ROACH | R04CH | COAT | C04T |
EAGLE | 346L3 | LADYBUG | L4DYBU6 | PANTS | P4NT5 |
DOVE | D0V3 | MAGGOT | M4660T | SCARF | 5C4RF |
Family Members | Musical Instruments | Fruit | |||
Word | LEET | Word | LEET | Word | LEET |
COUSIN | C0U51N | BAGPIPE | B46P1P3 | GUAVA | GU4V4 |
GRANDMA | 6R4NDM4 | GUITAR | 6U1T4R | LIME | L1M3 |
NIECE | N13C3 | HARP | H4RP | PEAR | P34R |
NEPHEW | N3PH3W | OBOE | 0B03 | BANANA | B4N4N4 |
WIFE | W1F3 | CYMBALS | CYMB4L5 | ORANGE | 0R4N63 |
MOTHER | M0TH3R | CELLO | C3LL0 | APPLE | 4PPL3 |
UNCLE | UNC13 | VIOLIN | V10L1N | APRICOT | 4PR1C0T |
DAD | D4D | BASS | B455 | BERRY | 83RRY |
SISTER | 515T3R | VIOLA | V1OL4 | NECTARINE | N3CT4RIN3 |
SIBLING | 51BL1N6 | ORGAN | 0R64N | MANGO | M4NG0 |
SPOUSE | 5P0U53 | PICCOLO | P1CC0L0 | GRAPE | GR4P3 |
PARENT | P4R3NT | BASSOON | 845500N | FIG | FI6 |
AUNT | 4UNT | TUBA | TU84 | RAISIN | R41S1N |
FATHER | F4TH3R | BANJO | B4NJ0 | AVACADO | 4V4C4D0 |
SON | S0N | CLARINET | CL4R1N3T | LEMON | L3M0N |
GRANDPA | 6R4NDP4 | PIANO | P14N0 | MELON | M3L0N |
BROTHER | BR0TH3R | FIDDLE | F1DDL3 | PEACH | P34CH |
HUSBAND | HU5B4ND | BUGLE | BU6L3 | PAPAYA | P4P4Y4 |
Vegetables | Trees/Flowers/Plants | Furniture | |||
Word | LEET | Word | LEET | Word | LEET |
CELERY | C3L3RY | GRASS | 6R455 | DRAWER | DR4W3R |
BEANS | B34N5 | CEDAR | C3D4R | DRESSER | DR3553R |
POTATO | P0T4T0 | ROSE | R053 | CHAIR | CH41R |
LETTUCE | L3TTUC3 | ELM | 3LM | TABLE | T4BL3 |
ONION | 0N10N | ASH | 45H | CABINET | C4B1N3T |
PEPPER | P3PP3R | BEECH | B33CH | ARMOIRE | 4RM01R |
CARROT | C4RR0T | MAPLE | M4PL3 | BED | B3D |
SPINACH | 5P1N4CH | BUCKEYE | BUCK3Y3 | ROCKER | R0CK3R |
BEETS | B33T5 | GINKO | 61NK0 | STOOL | 5T00L |
RADISH | R4D15H | SPRUCE | 5PRUC3 | DESK | D35K |
LEEK | L33K | REDWOOD | R3DW00D | RACK | R4CK |
CABBAGE | C4BB463 | FERN | F3RN | SHELF | 5H3LF |
PEA | P34 | OAK | 04K | VANITY | V4N1TY |
GARLIC | 64RL1C | LILAC | L1L4C | STAND | 5T4ND |
SPROUTS | 5PR0UT5 | CYPRESS | CYPR355 | CHEST | CH35T |
YAM | Y4M | VIOLET | V10L3T | BENCH | 83NCH |
PEANUT | P34NUT | PEONY | P30NY | SOFA | 50F4 |
SQUASH | 5QU45H | PINE | P1N3 | LAMP | L4MP |
Occupations | Money | Room/Place in a House | |||
Word | LEET | Word | LEET | Word | LEET |
ACTOR | 4CT0R | EURO | 3UR0 | PATIO | P4T10 |
JANITOR | J4N1T0R | BILL | 81LL | DOORWAY | D00RW4Y |
BANKER | B4NK3R | DOLLAR | D0LL4R | PARLOR | P4RL0R |
LAWYER | L4WY3R | BUCK | 8UCK | GARAGE | G4R46E |
CLERK | CL3RK | LOAN | L04N | CELLAR | C3LL4R |
ARTIST | 4RT15T | CASH | C45H | KITCHEN | K1TCH3N |
FARMER | F4RM3R | DIME | D1M3 | ATRIUM | 4TR1UM |
JUDGE | JUD63 | COIN | C01N | BALCONY | 84LC0NY |
BARBER | B4RB3R | DEBT | D38T | HALLWAY | H4LLW4Y |
TEACHER | T34CH3R | QUARTER | QU4RT3R | PANTRY | P4NTRY |
MANAGER | M4N463R | YEN | Y3N | FOYER | F0Y3R |
CHEF | CH3F | CREDIT | CR3D1T | ATTIC | 4TT1C |
BAKER | B4K3R | NICKEL | N1CK3L | DEN | D3N |
SAILOR | S41L0R | ACCOUNT | 4CC0UNT | BEDROOM | B3DR00M |
WRITER | WR1T3R | PAY | P4Y | STEPS | 5T3P5 |
THERAPIST | TH3R4PI5T | CHECK | CH3CK | DECK | D3CK |
COACH | C04CH | PENNY | P3NNY | CLOSET | CL0S3T |
FIREMAN | F1R3M4N | PESO | P3S0 | STAIRS | ST4IR5 |
Cooking Tools/Appliances | Geographical Features | Weather | |||
Word | LEET | Word | LEET | Word | LEET |
PEELER | P33L3R | OCEAN | 0C34N | SLEET | 5L33T |
FRIDGE | FR1D63 | STREAM | 5TR34M | DRIZZLE | DR1ZZL3 |
SPATULA | 5P4TUL4 | FOREST | F0R35T | RAINBOW | R41N80W |
GRILL | 6R1LL | DESERT | D353RT | TWISTER | TW15T3R |
TOASTER | T045T3R | BEACH | B34CH | HAIL | H41L |
KNIFE | KN1F3 | SEA | S34 | MIST | M15T |
FAUCET | F4UC3T | CANYON | C4NY0N | CYCLONE | CYC10N3 |
BASTER | B45T3R | BROOK | 8R00K | GALE | 64L3 |
SINK | 51NK | CAVE | C4V3 | BREEZE | BR33Z3 |
WHISK | WH15K | MEADOW | M34D0W | FROST | FR05T |
OVEN | 0V3N | CANAL | C4N4L | RAIN | R41N |
BLENDER | 8L3ND3R | LAKE | L4K3 | TORNADO | T0RN4D0 |
MIXER | M1X3R | VALLEY | V4LL3Y | SKY | 5KY |
GRATER | 6R4T3R | SWAMP | 5W4MP | STORM | 5TR0RM |
PAN | P4N | CREEK | CR33K | FOGGY | F066Y |
STOVE | 5T0V3 | ISLAND | 15L4ND | SNOW | 5N0W |
TEAPOT | T34P0T | BAY | 84Y | HAZY | H4ZY |
LADLE | L4DL3 | RIVER | R1V3R | SHOWER | 5H0W3R |
Colors | Fish | Emotion/Expression/Feeling | |||
Word | LEET | Word | LEET | Word | LEET |
BLACK | 8L4CK | EEL | 33L | FEAR | F34R |
WHITE | WH1T3 | SOLE | 50L3 | TERROR | T3RR0R |
TAN | T4N | PERCH | P3RCH | FURIOUS | FUR10U5 |
SILVER | 51LV3R | BASS | B455 | UPSET | UP53T |
YELLOW | Y3LL0W | HALIBUT | H4L18UT | SAD | 54D |
CYAN | CY4N | GROUPER | 6R0UP3R | DESIRE | D351R3 |
GRAY | 6R4Y | SALMON | 54LM0N | ANGER | 4N63R |
INDIGO | 1ND160 | SHARK | 5H4RK | DISGUST | D156U5T |
MAGENTA | M463NT4 | CARP | C4RP | BLISS | BL155 |
VIOLET | V10L3T | HADDOCK | H4DD0CK | LUST | LU5T |
BRONZE | 8R0NZ3 | TUNA | TUN4 | HOPE | H0P3 |
BLUE | 8LU3 | SARDINE | 54RD1N3 | ANXIETY | 4NX13TY |
GREEN | 6R33N | MARLIN | M4RL1N | GREED | 6R33D |
RED | R3D | WALLEYE | W4LL3Y3 | HAPPY | H4PPY |
MAROON | M4R00N | SQUID | 5QU1D | MAD | M4D |
BROWN | 8R0WN | HERRING | H3RR1N6 | GRUMPY | 6RUMPY |
GOLD | 60LD | CATFISH | C4TF15H | ENVY | 3NVY |
BEIGE | B31G3 | SNAPPER | 5N4PP3R | GLAD | 6L4D |
Sports | |
Word | LEET |
TRACK | TR4CT |
BOXING | B0X1N6 |
SURFING | 5URF1N6 |
FENCING | F3NC1N6 |
TENNIS | T3NN15 |
CYCLING | CYCL1N6 |
GOLF | 60LF |
RUGBY | RU68Y |
RACING | R4C1N6 |
SKIING | 5K11N6 |
HOCKEY | H0CK3Y |
ROWING | R0W1N6 |
CROQUET | CR0QU3T |
SOCCER | 50CC3R |
ARCHERY | 4RCH3RY |
KARATE | K4R4T3 |
BOWLING | 80WL1N6 |
DIVING | D1V1N6 |
Rights and permissions
About this article
Cite this article
Lien, MC., Allen, P. & Martin, N. Processing visual words with numbers: Electrophysiological evidence for semantic activation. Psychon Bull Rev 21, 1056–1066 (2014). https://doi.org/10.3758/s13423-014-0581-x
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13423-014-0581-x