
Abstract
The role of inhibition in the task-switching process has received increased empirical and theoretical attention in the literature on cognitive control. Many accounts have suggested that inhibition occurs when a conflict must be resolved—for example, when a target stimulus contains features of more than one task. In the two experiments reported here, we used variants of backward inhibition, or N – 2 repetition, designs to examine (1) whether inhibition occurs in the absence of conflict at the stimulus or response level, (2) when in the task-switching process such inhibition may occur, and (3) the potential consequences of inhibition. In Experiment 1, we demonstrate that neither stimulus- nor response-level conflict is necessary for inhibition to occur, while the results of Experiment 2 suggest that inhibition may be associated with a reduction of proactive interference (PI) from a previously performed task. Evidence of inhibition and the reduction of PI both occurred at the task-set level. However, inhibition of specific stimulus values can also occur, but this is clearly separable from task-set inhibition. Both experiments also provided evidence that task-set inhibition can be applied at the time of the new task cue, as opposed to at the onset of the target or at the response stage of the trial. Taken together, the results from these experiments provide insight into when and where in the task-switching process inhibition may occur, as well as into the potential functional benefits that inhibition of task sets may provide.
Similar content being viewed by others

The act of switching among multiple tasks is very familiar to most of us, and we often have an intuitive sense of the costs of such switching. If one stops reading in order to receive a phone call, most likely on a very different topic, this task switch seems to require some degree of cognitive control, but the effort required to return to reading may seem even greater. The consequences of switching and the cognitive control mechanisms underlying task-switching processes have been the focus of much recent research. In the experiments reported here, we addressed these issues, focusing in particular on when and how inhibitory processes may operate using variations of the backward inhibition (BI) paradigm originally described by Mayr and Keele (2000). We begin with a brief discussion of the costs and interference patterns typically found during task switching and the conditions under which inhibition has been demonstrated. Two experiments are then described that specifically explored (1) whether either stimulus or response competition is necessary to trigger inhibitory processes, (2) what the consequences of inhibition might be with respect to interference from the previous task, and (3) whether inhibition operates at the stimulus or task-set level.
Most research in this area has demonstrated that switching among multiple tasks results in time and accuracy costs as compared to repeating the same task. This switch cost has been found across a wide variety of experimental designs, although theoretical accounts of the sources and components of the switch cost vary (for reviews, see Kiesel et al., 2010; Vandierendonck, Liefooghe, & Verbruggen, 2010). While much work on task switching has focused on how a new task is activated, reconfigured, or retrieved from memory (e.g., Mayr & Kliegl, 2000; Rogers & Monsell, 1995), the process of preparing a new task cannot account for all aspects of the observed response time (RT) and accuracy patterns. Instead, some have suggested that interference from the previous task and/or inhibition of that task must also be considered (e.g., Kiesel et al., 2010; Monsell, 2003). Interference accounts suggest that the influence of earlier trials persists into later trials or affects new task-set retrieval, and thus interferes proactively with performance of the new task (e.g., Allport, Styles, & Hsieh, 1994; Wylie & Allport, 2000). Importantly, such accounts imply that old task sets are not actively inhibited, allowing proactive interference to occur.
In contrast, some accounts posit that when one switches to a new task, the old task is actively inhibited. The most commonly cited evidence of inhibition comes from the finding that it takes longer to switch back to a task that was recently performed (e.g., ABA) than to one that was not (e.g., CBA). This measure of inhibition, also referred to as the N – 2 repetition cost, has been investigated in a variety task-switching designs (see Koch, Gade, Schuch, & Philipp, 2010, for a review). Such an inhibitory process would seem to imply that recent tasks, having been inhibited, should not produce interference on subsequent trials.
Inhibition generally has been assumed to be useful for resolving conflict between task sets, either at the stimulus or the response level. For bivalent targets containing features relevant to two tasks (e.g., a red triangle), inhibition may serve to reduce perceptual interference from the irrelevant feature. Similarly, the “red” and “triangle” features could produce response competition that must be resolved. While it is commonly accepted that such conflict and conflict resolution processes may increase switch costs, some have argued that response- or stimulus-level conflict is a necessary condition to produce inhibition effects (e.g., Koch et al., 2010; Mayr & Keele, 2000), or even switch costs (e.g., Meiran, 2000).
If inhibition does operate during task switching, it is important to understand what factors modulate the inhibitory processes. For example, the amount of response competition in the experiment as a whole may influence whether and how much inhibition occurs on a given trial (e.g., Brown, Reynolds, & Braver, 2007; Gade & Koch, 2007; Lien, Ruthruff, & Kuhns, 2006). It has also been suggested that inhibition may be influenced by task demands. For example, the presence of bivalent stimuli in a stimulus set may trigger stronger inhibitory processes, due to increased conflict overall (Lien et al., 2006). Such an incentive may also be influenced by how frequently task switches occur, so that eliminating task repeats in the stimulus set could eliminate inhibition (Philipp & Koch, 2006). By maximizing the incentive for task-set inhibition, Kuhns, Lien, and Ruthruff (2007) demonstrated that under some conditions inhibition may be applied proactively, or prior to the onset of the new target stimulus. This account stands in contrast to models that have suggested that inhibition is applied reactively, in response to immediate conflict, although proactive and reactive inhibitory processes need not be mutually exclusive.
The two experiments reported here were designed to explore the necessary conditions for task-set inhibition, as well as the nature of such inhibitory processes. In Experiment 1, we directly addressed the assumption that conflict, in the form of stimulus or response multivalency, is necessary for inhibition to occur. Experiment 2 provided a richer picture of the inhibitory processes by measuring both inhibition and its potential benefits in terms of the control of proactive interference. In both experiments, we considered what type of information is inhibited and when in the task structure inhibition may occur.
Experiment 1
If an inhibitory mechanism is associated with disengaging from one task when switching to another, one might ask whether such inhibition is a necessary process and what function such a process might serve. Several experiments have attempted to address these questions by focusing on whether the amount of inhibition found is linked to the presence or the degree of response competition on a given trial (see Koch et al., 2010). Schuch and Koch (2003), for example, found no evidence of BI after trials that did not require a response, suggesting that BI operates reactively, at the point of response selection; however, Kuhns et al. (2007) showed that if the difficulty of task switching is increased through the inclusion of multivalent stimuli and responses in the experimental design, inhibition then appears to occur proactively. Similarly, Mayr and Keele (2000, note 2) considered the possibilities that inhibition develops over time as a result of competition between task sets and responses that is triggered by multivalent stimuli, and that little or no inhibition should be expected if conflict resolution is not necessary. However, seemingly contradictory to this assertion, they demonstrated a small but significant BI effect when no response conflict was present and only one dimension of the stimulus target afforded a response (Mayr & Keele, 2000, Exp. 3). Thus, response conflict within a target may not be a necessary condition for inhibition to occur. Finally, Lien et al. (2006) also attempted to determine whether inhibition depends on conflict. They did not look specifically at BI effects, but used a mix of bivalent and univalent stimuli in order to manipulate the incentive for inhibition, concluding that inhibition is not sensitive to this type of context effect and that task-set inhibition may be weak or easily overcome.
The most extreme test of the role of conflict resolution in triggering inhibitory processes is to eliminate conflict at both the stimulus and response levels. Accordingly, we used univalent stimuli with features from only one task, as well as univalent response mapping, with each response button associated with only one stimulus feature. Thus, conflict was not present either in the stimulus or from competing response activation at any point in this experiment. If inhibition is triggered by conflict resolution processes, we should thus find no evidence of BI. However, if inhibition occurs as a result of disengaging from one task or preparing another, we should find significant BI.
Method
Apparatus and participants
The experiment was run on E-Prime version 1.0 software and displayed on an 18-in. monitor. Responses were made on a custom-built Psychological Software Tools serial response box expansion kit with six button switches. A group of 36 college-aged participants (21 female, 15 male) were recruited from undergraduate psychology classes and received extra course credit; however, the data from one participant were lost due to computer error, so the analyses are based on N = 35.
Tasks and stimuli
For the experiment we utilized three tasks, each with two associated features: red/green for the color task, horizontal/vertical lines for line orientation, and a triangle or circle for shape. The relevant task for the upcoming trial was explicitly cued in advance of the target with the word COLOR, LINE, or SHAPE. Each stimulus had only one dimension and did not contain features from either of the other two tasks (see Table 1). The RTs from the onset of the target stimulus to the buttonpress and accuracy were recorded.
Procedure and design
Participants completed one training block of 20 trials for each task, with no task switches (i.e., 20 color, 20 line, and 20 shape trials), and a final training block of 40 trials that consisted of all three tasks with the task repeat and switch conditions randomly intermixed. The orders in which the participants saw the three tasks in the no-switch training blocks were counterbalanced across participants. Each target type was associated with one of six buttons on the response box, with each response key located equidistant from a center finger rest pad (see Fig. 1). The locations of the responses were rotated across participants to counterbalance the target–response key mappings. Participants were instructed to respond using the index finger of their preferred hand and to return their finger to the central rest pad after each response.

The response box used for both experiments. The small black circles represent the response keys, while the light-gray center circle represents the finger rest pad. The response key assignments for each task were counterbalanced across subjects. In this example, for the color task key labels (lower left), the gray shade represents the color green and black represents the color red
After training, the participants completed ten experimental blocks of 50 trials each. They could take a brief, self-timed break between blocks, and cumulative RTs and accuracy were reported to the participants after each block to motivate them to respond quickly and accurately. The three tasks were presented in a pseudorandom order, and each of the ten blocks contained a unique sequence of trials, such that neither the task, the target, nor the cue–target interval (CTI) of the next trial could be predicted on the basis of previous sequences. The task cue was presented 500 ms after the response to the previous trial and appeared either 500 ms (short) or 1,000 ms (long) before the onset of the target stimulus, creating two CTIs. The cue disappeared at the onset of the target stimulus.
On approximately one-third of the trials, participants repeated the same task that they had performed on the previous trial (task repeats), while they switched to a new task on approximately two-thirds of the trials (task switches). Half of the task switches were BI conditions (N – 2 repetitions) that required the participants to switch back to the task that had been performed on trial N – 2 (e.g., ABA), while the other half of the switches were not (e.g., CBA). All of the variables were counterbalanced across CTIs.
Results
Errors, posterror trials, and trials with RTs less than 200 or greater than 2,000 ms were excluded from the RT analyses. Because BI required that each trial be coded relative to trial N – 2, the first two trials of each block were also excluded from the BI analyses. The same procedures (other than error removal) were used for the accuracy analyses. Within-subjects median RTs and accuracy were then computed for each condition. Accuracy was generally very high (all condition accuracy means were greater than 95 %), and where significant accuracy effects were found, conditions of greater accuracy were associated with faster RTs. There was no evidence of a speed–accuracy trade-off; thus, only the RT analyses are reported.
Overall analysis
A 2 (task switch) × 2 (CTI) ANOVA was conducted on the median RT data to measure the effects of switch cost and preparation interval. The main effects were significant for task switch [F(1, 34) = 109.37, p < .01, η 2p = .76] and CTI [F(1, 34) = 143.32, p < .01, η 2p = .81], with faster RTs for task repeats than for switches and for long than for short CTIs. The Task Switch × CTI interaction was also significant [F(1, 34) = 60.07, p < .01, η 2p = .64], with a smaller switch cost at the long (45-ms cost) than at the short (99-ms cost) CTI.
BI analysis
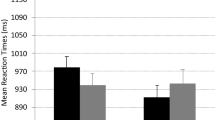
A 2 (BI) × 2 (CTI) ANOVA was conducted on the task switch trials, revealing a main effect of BI [F(1, 34) = 21.28, p < .01, η 2p = .39], with longer RTs when the most recently disengaged task was reactivated (see Table 2). The main effect of CTI was also significant [F(1, 34) = 169.99, p < .01, η 2p = .83], with faster RTs for the long CTI; however, CTI did not interact with BI [F(1, 34) = 2.45, p = .13].
A post-hoc dependent-samples t test was conducted to determine whether the BI effect occurred for all N – 2 task repeats or only when the specific stimulus value (e.g., a triangle) occurred on both trials N – 2 and N. The difference was not significant, t(34) = 0.35, p = .73, suggesting that BI occurs at the task-set level.
Discussion
We found significant RT costs for switching tasks, with the cost reduced but not eliminated at the longer CTI, a pattern similar to typical results found with multivalent stimuli. Thus, switch costs and the benefit of a preparation interval occurred even when using only univalent stimuli and responses. Most importantly, we also observed significant inhibition when univalent stimuli and response mappings were used. The presence of significant BI when only univalent stimuli were present demonstrates that conflict between multiple dimensions of the target stimulus is not a necessary condition for inhibition to occur, nor is response competition within the target.Footnote 1 Inhibition, then, appears to occur not specifically in reaction to previous or potential interference within an upcoming target, but rather as a more general process of disengaging from or preparing a task. This inhibitory effect also occurred at the task-set and not at the stimulus-value level, suggesting that such inhibition is proactive, not reactive, in nature (Kuhns et al., 2007). In general, proactive inhibition might be expected when the incentive for inhibition is high—that is, with increased levels of interference and when the task switches on every trial. The results of the present experiment are surprising because of the minimized interference within a trial and because of the inclusion of task repeat trials, which should reduce the incentive for proactive inhibition. When every trial requires a switch from the previous task, proactive inhibition may speed the switch process; however, when task repeat trials are included (on one-third of the trials, in the present case), that strategy would be counterproductive. Indeed, Philipp and Koch (2006) reported that including task repeat trials decreased or eliminated N – 2 repetition costs. Nonetheless, in the present study we found significant BI under such conditions, suggesting that inhibition may be tied to task disengagement and/or to new task preparation rather than to conflict resolution.
Experiment 2
Experiment 1 demonstrated that, in contrast to previous reports, inhibition in the form of N – 2 repetition costs can be found under conditions that offer no incentive for inhibition: The presence of task repeat trials should discourage a prepare-to-switch strategy, and there was minimal competition within a trial, due to the use of univalent targets and responses. By definition, this inhibition cannot be considered to be reactive, in the sense of being triggered by conflict present within the target. The inhibition, then, appears to be proactive, aiding new task preparation.
Previous reports supporting the notion of proactive inhibition have suggested that such inhibitory processes may serve to reduce interference from abandoned tasks (e.g., Hübner, Dreisbach, Haider, & Kluwe, 2003; Kuhns et al., 2007). There are several possible ways in which the previous trial could interfere with the current trial: The relevant task set (e.g., all stimuli and responses related to shape) or the specific stimulus value (e.g., a triangle) on trial N – 1 could produce proactive interference if it had occurred as the irrelevant task or value on trial N. Similarly, the irrelevant task set or a specific stimulus value on trial N – 1 could produce negative priming if that task or value was ignored on trial N – 1 but is relevant on trial N. Kuhns et al. (2007), using a flanker paradigm, argued that proactive inhibition controlled interference at the task-set level. However, theirs and other studies (i.e., Mayr & Keele, 2000) were designed specifically to exclude the possibility of stimulus-specific effects.
In principle, both reactive and proactive inhibition could operate within the same task. Even when there is no immediate conflict, proactive inhibition may occur as a task is disengaged, in order to facilitate the switch process. Such inhibition may have the benefit of reducing interference from the abandoned task under some conditions, but it also carries a potential cost if an inhibited task must be reactivated. In addition, a reactive inhibition process that occurs when features from two or more tasks occur simultaneously may be required in order to resolve within-stimulus competition.
In Experiment 2, we explored both the potential sources of interference from trial N – 1 and the consequences of inhibition with respect to those carryover effects. In this experiment, the targets were bivalent, which allowed for the measurement of potential N – 1 interference at both the stimulus-value and the task-set levels. Specifically, we were interested in whether the proactive inhibition found in Experiment 1 served to reduce interference from the previous trial.
Method
Apparatus and participants
The apparatus and participant recruitment were the same as were described for the previous experiment, with 36 (17 female, 19 male) new participants.
Tasks and stimuli
The same three tasks were used as in Experiment 1, but with bivalent stimuli (see Table 1): each stimulus had one relevant task feature and one irrelevant feature from one of the other two tasks. The use of bivalent stimuli with the three tasks allowed us to compare the amount of interference caused by stimuli related to the previous task set with that caused by stimuli associated with a task that had not been performed as recently. All possible combinations of the tasks were used, with each task appearing equally often as the relevant or irrelevant task.
Procedure and design
The training procedure, response box, and experimental procedure, including the task cues and CTIs (500 and 1,000 ms) were the same as in Experiment 1. The task was again repeated on one-third of the trials, and was switched on the other two-thirds. Half of the task switch trials required reactivating the task from trial N – 2 (BI), while the other half did not (no BI).
Two additional variables allowed us to measure potential interference from the previous trial (N – 1), which could occur in the form of either proactive interference (PI) or negative priming (NP). In the PI conditions, the task performed on trial N – 1 served as the distractor on trial N (e.g., COLOR/line➔SHAPE/color, with the repeated feature set in boldface). In the non-PI conditions, the target task from trial N – 1 did not appear in the subsequent trial (e.g., COLOR/line➔SHAPE/line). In the NP condition, the task ignored on trial N – 1 occurred as the relevant task on trial N (e.g., COLOR/shape➔SHAPE/line), but not in the non-NP conditions (e.g., COLOR/shape➔LINE/shape). The BI factor and the two N – 1 interference variables were crossed within each CTI. Finally, stimulus-value-level interference conditions were nested within each task-set-level condition to determine whether the observed interference was associated with a repetition of the task set in general (e.g., color) or with the specific stimulus value (e.g., red).
Results
The data-trimming procedures were the same as those described for Experiment 1. Accuracy rates were high, with no evidence of a speed–accuracy trade-off.
Overall analysis
A 2 (task switch) × 2 (CTI) ANOVA was conducted to measure the overall switch cost and the preparation effects. The task switch cost was reliable [F(1, 35) = 87.41, p < .05, η 2p = .71], as was the main effect of CTI [F(1, 35) = 173.40, p < .01, η 2p = .83], with faster RTs for the long interval. Task switch and CTI interacted significantly [F(1, 35) = 45.00, p < .01, η 2p = .56], with larger switch costs at the short CTI (106 ms) than at the long (37 ms).
BI analyses
Potential BI effects were examined through a 2 (BI) × 2 (CTI) ANOVA of the task switch trials (see Table 2). The effect of BI was significant [F(1, 35) = 4.65, p < .05, η 2p = .12], with N – 2 repetitions having longer RTs than non-(N – 2) repetitions. The effect of CTI was also significant [F(1, 35) = 139.96, p < .01, η 2p = .80], with faster RTs at the long CTI, but CTI did not interact with BI [F(1, 35) = 1.71, p = .20].
An additional analysis was conducted to determine what information was inhibited. As in Experiment 1, the dependent-samples t test was not significant, t(35) = 1.88, p = .07, suggesting that N – 2 repetition costs occurred whenever the N – 2 task repeated, regardless of whether or not the specific stimulus value was repeated.
Interference analyses
Task-set-level interference
In order to examine the two possible forms of interference, a 2 (PI) × 2 (NP) × 2 (CTI) ANOVA was conducted within the task switch trials. The PI effect was significant [F(1, 35) = 9.67, p < .01, η 2p = .22], showing a reduction in interference consistent with inhibition; that is, RTs were faster when the relevant task on N – 1 appeared as an irrelevant task on trial N than they were on trials in which it did not (see Table 3). The effect of NP was also significant [F(1, 35) = 8.15, p < .05, η 2p = .19], with NP trials being slowed as compared to non-NP trials. Finally, the CTI effect was significant [F(1, 35) = 139.85, p < .01, η 2p = .80], with short CTIs resulting in longer RTs (M = 713 ms, SD = 17.8) than the long intervals (M = 621 ms, SD = 18.9). PI and NP did not interact with each other or with CTI.
Feature-level interference
To evaluate interference due to the repetition of a specific stimulus value across sequential trials, we conducted dependent-samples t tests for stimulus-specific repetitions versus nonrepetitions for both PI and NP. For PI, stimulus-value repetition was not significant, t(35) = 0.56, p = .58, suggesting that if the task responded to on trial N – 1 was used as the distractor on trial N, it did not matter whether the specific stimulus value (e.g., “red”) was repeated (e.g., Rv➔Tr; M = 657 ms, SD = 17.0) or not (e.g., Rv➔Tg; M = 653 ms, SD = 18.2); rather, it appears that the PI effect occurs at the task-set level.
The same dependent-samples t test comparing stimulus-value repetitions versus nonrepetitions for NP was significant [t(35) = 2.25, p < .05, d = 0.37], reflecting a significant cost if the specific stimulus (e.g., “red”) that was ignored on trial N – 1 became the relevant response (e.g., Tr➔Rv; M = 684 ms, SD = 20.2), as compared to when the same task but not the specific value became relevant on the subsequent trial (e.g., Tr➔Gv; M = 665 ms, SD = 18.4). This finding suggests that NP interference, unlike PI, is likely driven by reactive inhibitory processes at the level of the stimulus, not the task set.
Discussion
This experiment sheds additional light on the nature and the consequences of the inhibitory processes involved in task switching. First, significant BI effects were found at the task-set level that appear to be “immune” to CTI effects. As in Experiment 1, one-third of the trials were task repeat trials, discouraging a strategy of preparing to switch after every response. These results suggest that inhibition occurs when the cue indicating a task switch appears, consistent with Hübner et al. (2003).
These results are also consistent with Kuhns et al.’s (2007) argument that BI reflects a proactive form of inhibition that reduces interference in cases in which the N – 1 task occurs as the irrelevant task on trial N. While Kuhns et al. employed a flanker task with distractors separate from the response-relevant target, in the present experiment we used bivalent targets, integrating the relevant and irrelevant task features. Because BI is a measure of N – 2 repetition costs, and the reduction of PI is based on task repetition from trial N – 1, it is not possible to measure both on the same trial. However, our results show similarities in the patterns of BI and interference reduction: Both occur at the task-set level, and neither is sensitive to CTI manipulations. Whether or not BI is the cause, this reduction of PI has important implications for models of task switching. Some accounts have suggested that either persisting activation of previously relevant tasks or episodic retrieval of prior S–R mappings may produce switch costs due to interference from trial N – 1 (e.g., Allport et al., 1994; Koch & Allport, 2006; Wylie & Allport, 2000). These results suggest that proactive inhibition typically reduces such interference from the previous trial.
Finally, these results demonstrate a clear distinction between the proactive and reactive inhibition processes suggested by Kuhns et al. (2007). Proactive inhibition processes were clearly task-set-level effects. In contrast, a significant negative-priming effect was found at the stimulus-value level rather than at the task-set level. This suggests that reactive inhibition occurs when a target with multiple task features must be responded to; however, this form of inhibition operates on the level of the specific stimulus value and is separable from task-set-level proactive inhibition.
General discussion
The focus of this work was on the nature and potential consequences of inhibitory mechanisms in the task-switching process. Retrieving task information from memory and preparing that information in advance of a target stimulus takes time but in principle can be accomplished through facilitation processes, without the inhibition of previous tasks or distracting information. However, an extensive body of work has suggested that, at least under some circumstances, inhibitory processes contribute to the switch process, if not to switch costs per se (e.g., Koch et al., 2010). In these experiments, we found that BI can occur with univalent stimuli and response mappings (Exp. 1) and that proactive interference from the previous task is reduced in a manner consistent with this type of inhibition (Exp. 2).
An underlying assumption in this literature has been that perceptual or response competition is necessary to trigger inhibition (e.g., Gade & Koch, 2005, 2007; Koch et al. 2010; Mayr & Keele, 2000; Schuch & Koch, 2003). The results of Experiment 1 clearly demonstrate that task competition in the form of either (1) a stimulus with features of more than one task or (2) shared response mappings across multiple tasks is not a necessary condition for inhibition, as measured in terms of BI. These findings may be especially surprising, given that none of the usual conditions were present that might increase the incentive to adopt an inhibitory strategy in general. Univalent response mappings were used in both experiments, a factor that should make the strategic use of inhibition less likely (Kuhns et al., 2007). Additionally, many task-switching studies have included only switch trials in the design, possibly encouraging a strategy of preparing for the next trial (and task) by inhibiting the last one. In the experiments reported here, approximately one-third of the trials were task repeats; even so, significant BI was found.
The results of these experiments suggest that a proactive inhibition process, such as the one described by Kuhns et al. (2007), may be the source of the BI effect. But if neither the targets nor the response mappings elicit task competition, what is being inhibited? The data from both experiments indicate that inhibition occurred at the task-set level rather than at the specific stimulus-value level. In the absence of conflict within a trial, it seems likely that disengaging from the previous task or preparing the new task is in itself sufficient to trigger inhibition of the abandoned task. Indeed, these results seem consistent with the suggestion made by Houghton, Pritchard, and Grange (2009; see also Grange & Houghton, 2010). They proposed that the cue is used to retrieve the current task set from long-term memory and that linking the cue to task information makes demands on working memory (WM). Inhibition of the previous task set at the point of the next cue will effectively clear out WM for the new-cue processing and task-set retrieval. Inhibition of the entire task set (rather than of the features of the previous target) in the absence of any target-based stimulus or response competition makes sense, in the context of intertrial WM demands.
Experiment 2 showed evidence of both BI and a reduction of proactive interference from trial N – 1 that is consistent with inhibition of the previous task. Although explanations of switch costs have been based on carryover effects from the previous trial (see Vandierendonk et al., 2010), a logical implication of the BI evidence here is that there should be less interference from the abandoned, and therefore inhibited, task from the previous trial. Using flanker task designs, Hübner et al. (2003) and Kuhns et al. (2007) also found evidence of reduced PI, but only under some conditions. The nature of the cuing process was an important factor in Hübner et al.’s study, such that interference control did not occur with no cue or when a switch was expected but the specific task was not known. Kuhns et al. reported that no reduction of PI was found when the response mapping was univalent and the target occurred predictably between two flankers; however, reduced interference was found with a combination of multivalent responses and unpredictable target locations, which the authors believed more closely resembled the type of bivalent targets typically used in task-switching studies. On the basis of our first experiment, we suspect that a predictable target location was the crucial factor determining the PI effect.
It is noteworthy that both BI and PI reduction occurred at the task-set level, which would be expected if this interference control was the result of the inhibitory processes measured by BI. In contrast, we also found significant interference effects from trial N – 1 in the form of negative priming, but not at the task-set level: RTs were slower if the current task was ignored on the previous trial, but only when the ignored feature (“triangle”) on trial N – 1 matched the target feature on trial N.
Inhibitory processes seem to occur at several different levels and to have a variety of consequences. When a multivalent stimulus is being processed and a separation of relevant and irrelevant features or responses is necessary, inhibitory processes are triggered that are specific to the irrelevant features and responses. The results of such inhibition may carry over to the next trial. However, that process is clearly distinct from inhibitory processes that appear to be based on cue presentation and new task preparation—proactive inhibition, in Kuhns et al.’s (2007) terms. Such processes may serve the function of clearing WM of its previous contents (Houghton et al., 2009), as a result of which the entire task set is inhibited. As we demonstrated in the present experiments, such inhibition is not dependent on multivalent stimuli or response mappings; it may aid task switching by reducing interference from the previous trial, but it can also produce costs if the task needs to be reengaged immediately.
Notes
Traditionally, a response mapping is deemed univalent when each target is associated with a different response key, as in the present case. An interesting consideration, however, is how distinct the motor responses must be in order to ensure that response competition is minimized. In these experiments, the same finger was used to press each of the six keys; is it possible that response competition was intrinsic to this situation? Similarly, does response competition occur if several fingers on one hand are used to make separate responses? The true independence of different types of responses is an important question, but we believe that the response procedure in the present case represents a univalent response mapping for several reasons. First, an analysis of costs based on how close the tasks were to each other on the response box showed no differences, suggesting that similarity of direction of movement did not affect performance. Second, other research from our lab involving univalent response mapping has used six fingers, one for each response key, and has demonstrated switch costs and BI effects consistent with the present results (Nelson & Friedrich, 2012).
References
Allport, A., Styles, E. A., & Hsieh, S. (1994). Shifting intentional set: Exploring the dynamic control of tasks. In C. Umiltà & M. Moscovitch (Eds.), Attention and performance XV (pp. 421–452). Hillsdale, NJ: Erlbaum.
Brown, J. W., Reynolds, J. R., & Braver, T. S. (2007). A computational model of fractionated conflict–control mechanisms in task-switching. Cognitive Psychology, 55, 37–85.
Gade, M., & Koch, I. (2005). Linking inhibition to activation in the control of task sequences. Psychonomic Bulletin & Review, 12, 530–534. doi:10.3758/BF03193800
Gade, M., & Koch, I. (2007). The influence of overlapping response sets on task inhibition. Memory & Cognition, 35, 603–609.
Grange, J. A., & Houghton, G. (2010). Heightened conflict in cue–target translation increases backward inhibition in set switching. Journal of Experimental Psychology: Learning, Memory, and Cognition, 36, 1003–1009.
Houghton, G., Pritchard, R., & Grange, J. A. (2009). The role of cue–target translation in backward inhibition of attentional set. Journal of Experimental Psychology: Learning, Memory, and Cognition, 35, 466–476.
Hübner, M., Dreisbach, G., Haider, H., & Kluwe, R. H. (2003). Backward inhibition as a means of sequential task-set control: Evidence for reduction of task competition. Journal of Experimental Psychology: Learning, Memory, and Cognition, 29, 289–297.
Kiesel, A., Steinhauser, M., Wendt, M., Falkenstein, M., Jost, K., Philipp, A. M., & Koch, I. (2010). Control and interference in task switching—A review. Psychological Bulletin, 136, 849–874. doi:10.1037/a0019842
Koch, I., & Allport, A. (2006). Cue-based preparation and stimulus-based priming of tasks in task switching. Memory & Cognition, 34, 433–444. doi:10.3758/BF03193420
Koch, I., Gade, M., Schuch, S., & Philipp, A. M. (2010). The role of inhibition in task switching: A review. Psychonomic Bulletin & Review, 17, 1–14. doi:10.3758/PBR.17.1.1
Kuhns, D., Lien, M.-C., & Ruthruff, E. (2007). Proactive versus reactive task-set inhibition in task switching: Evidence from flanker compatibility effects. Psychonomic Bulletin & Review, 14, 977–983.
Lien, M.-C., Ruthruff, E., & Kuhns, D. (2006). On the difficulty of task switching: Assessing the role of task-set inhibition. Psychonomic Bulletin & Review, 13, 530–535. doi:10.3758/BF03193881
Mayr, U., & Keele, S. W. (2000). Changing internal constraints on action: The role of backward inhibition. Journal of Experimental Psychology. General, 129, 4–26. doi:10.1037/0096-3445.129.1.4
Mayr, U., & Kliegl, R. (2000). Task-set switching and long-term memory retrieval. Journal of Experimental Psychology: Learning, Memory, and Cognition, 26, 1124–1140. doi:10.1037/0278-7393.26.5.1124
Meiran, N. (2000). Reconfiguration of stimulus task-sets and response task-sets during task-switching. In S. Monsell & J. S. Driver (Eds.), Control of cognitive processes: Attention and performance XVIII (pp. 377–400). Cambridge, MA: MIT Press.
Monsell, S. (2003). Task switching. Trends in Cognitive Sciences, 7, 134–140. doi:10.1016/S1364-6613(03)00028-7
Nelson, J., & Friedrich, F. (2012, April). Task variability and cognitive control. Poster presented at the Undergraduate Research Symposium, University of Utah, Salt Lake City, Utah.
Philipp, A. M., & Koch, I. (2006). Task inhibition and task repetition in task switching. European Journal of Cognitive Psychology, 18, 624–639.
Rogers, R. D., & Monsell, S. (1995). The cost of a predictable switch between simple cognitive tasks. Journal of Experimental Psychology. General, 124, 207–231. doi:10.1037/0096-3445.124.2.207
Schuch, S., & Koch, I. (2003). The role of response selection for inhibition of task sets in task shifting. Journal of Experimental Psychology. Human Perception and Performance, 29, 92–105. doi:10.1037/0096-1523.29.1.92
Vandierendonck, A., Liefooghe, B., & Verbruggen, F. (2010). Task switching: Interplay of reconfiguration and interference control. Psychological Bulletin, 136, 601–626. doi:10.1037/a0019791
Wylie, G., & Allport, A. (2000). Task switching and the measurement of “switch costs. Psychological Research, 63, 212–233. doi:10.1007/s004269900003
Author note
R.E.C. is now at the College of Arts and Sciences, Westminster College, Salt Lake City, Utah. This work is based on his doctoral dissertation.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Costa, R.E., Friedrich, F.J. Inhibition, interference, and conflict in task switching. Psychon Bull Rev 19, 1193–1201 (2012). https://doi.org/10.3758/s13423-012-0311-1
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13423-012-0311-1