
Abstract
Irrelevant salient distractors often capture attention, but given a sufficiently specific search template, these salient items no longer capture attention. In the present experiments, we investigated whether specific target templates are sufficient to resist capture, or whether experience with the salient distractors is also necessary. To test this hypothesis, observers completed four blocks of trials, each with a different-colored irrelevant singleton present on half of the trials. Color singletons captured attention early within a block, but after sufficient experience with the irrelevant singletons, those singletons no longer captured attention in the second halves of the blocks. This result suggests that to resist capture, a specific target template must be accompanied by experience-dependent attentional tuning to distractor properties.
Similar content being viewed by others

The world is filled with distractions, ranging from the abrupt appearance of an Internet pop-up ad to the sound of a coffee grinder at the local bistro. A primary purpose of attention is to restrict processing to items relevant to our current behavior and to minimize interference from irrelevant distractors. An important issue in the attentional-control literature has centered on the mechanisms of this restriction, with two rival hypotheses emerging to explain when attention is captured by distracting information. One account proposes that attention is stimulus-driven and that salient distractors capture attention irrespective of one’s goals or attentional set (Theeuwes, 1992, 2010). Another account proposes that attention is driven by one’s goals and that attentional capture is contingent on one’s attentional set; only distractors matching a current attentional set will capture attention (e.g., Folk, Remington, & Johnston, 1992). Against this backdrop, much of the past 20 years of research has focused on distinguishing stimulus-driven capture from contingent capture, with a focus on which of these two modes of selection is the “default” mode of attention (Kawahara, 2010).
Strong evidence for stimulus-driven capture has come from the additional-singleton paradigm (Theeuwes, 1992). In this task, observers search for a shape singleton among homogeneous distractors (e.g., a circle among diamonds) and report the orientation of a line that appears inside the target shape. On half of the trials, one of the distractors is a different color, making it a salient singleton distractor. Because the target is never the color singleton, an observer with perfect goal-driven control has no reason to attend to this additional singleton, and the presence of the singleton should not slow response times (RTs) to the target. However, the presence of an irrelevant color singleton does slow RTs to the target (see Theeuwes, 2010, for an extensive review). Because the color singleton is irrelevant to an observer’s goal of finding the shape singleton, slowed RTs when the color singleton is present can be interpreted as stimulus-driven attentional capture.
There are limits to stimulus-driven capture, however. One important retort to results from the additional-singleton paradigm came from Bacon and Egeth (1994), who demonstrated that a salient distractor would not capture attention when observers searched for a specific target, not simply a shape singleton target. Specifically, when searching for a target among heterogeneous distractors (e.g., a circle among squares, diamonds, and triangles), an irrelevant color singleton no longer captures attention. Bacon and Egeth reasoned that the specificity of the search template was critical in determining capture. When searching for a shape singleton, observers may adopt a nonspecific target configuration and search for any unique item—that is, any singleton. This so-called singleton detection mode (Pashler, 1988) would allow for fast detection of a singleton target, but would also render an observer vulnerable to capture by an irrelevant color singleton. Requiring observers to search for a target among heterogeneous distractors requires feature search mode, and searching for a specific feature (e.g., a circle) appears to eliminate capture because the color singleton distractor no longer matches the target properties (also see Folk et al., 1992).
Although Bacon and Egeth’s (1994) work highlighted the role of target specificity in attentional capture, subsequent work has demonstrated that attention is also affected by distractor attributes. For example, in priming of pop-out, Maljikovic and Nakayama (1994) not only demonstrated that observers were faster to respond to a target whose color repeated from the previous trial, they also found that observers were faster to discriminate the target when the distractor color repeated from the previous trial (see also Kristjánsson & Campana, 2010, for a review of priming of pop-out). This result suggests that attention is affected by distractor features (namely, color). Distractors also affect attention in contextual cuing (Chun & Jiang, 1998), in which repeated distractor positions improve visual search for a target. Finally, on the basis of separate electrophysiological signatures for attentional capture and distractor suppression, Sawaki and Luck (2010, 2011) recently proposed that salient distractors are detected, but attention is prevented from visiting the salient items by suppression of orienting to the distractor (see also Awh, Matsukura, & Serences, 2003, and Serences, Yantis, Culberson, & Awh, 2004, for discussions of distractor suppression).
Given this background, an outstanding question regarding attentional capture is how attention learns to suppress distractor items. One straightforward possibility is that a well-specified target template, such as that employed in feature search mode, dictates distractor suppression. Any item not matching the target template could be dismissed as a distractor. Support for this hypothesis comes from Leber, Kawahara, and Gabari (Leber et al. 2009), who found that if observers were trained to use a specific target template, color singletons failed to capture attention even after the color of the color singleton was switched. In contrast, we propose an alternative possibility: experience-dependent distractor rejection. On the basis of a wealth of findings that have indicated that attention is affected by recent experience, we propose that salient distractors must be encountered and must capture attention before they can be suppressed and rejected at some later time. In short, although a target template is likely to be important in configuring attention, attention may also need to tune itself to distractor properties in order to optimize selection.
To investigate these possibilities, observers performed a basic feature search task by searching for a circle among diamonds, squares, and triangles. Observers first learned the target template in a training block in which no color singleton distractor appeared. Following the training block, observers completed four test blocks in which a color singleton distractor appeared on half of the trials. Diverging from previous work, we presented a different-colored singleton distractor in each block. For example, the first block might contain a red color singleton, the second a purple color singleton, and so on. If a specific target template is sufficient for generating distractor suppression, the RTs to the target should be unaffected by the presence of a singleton distractor, replicating Bacon and Egeth’s (1994) well-known results. However, if efficient distractor rejection not only requires a specific target template, but also requires learning to reject specific distractors, we should observe attentional capture when observers initially experience a color singleton distractor. Specifically, in the first block in which a distractor appears, observers should initially be slower to respond when the color singleton distractor is present than when it is absent. Furthermore, our design allowed us to examine the generalization of any learned distractor rejection to novel color singletons in the following blocks. If exposure to any color singleton distractor is sufficient to learn distractor rejection, then we should observe no capture in Blocks 2, 3, and 4. Alternatively, if distractor-specific experience is required for distractor rejection, we should observe capture any time a new color singleton appears; specifically, we should observe capture at the beginnings of all of the blocks.
Experiment 1
Method
Participants
A group of 16 University of Iowa undergraduates participated for course credit. All reported having normal or corrected-to-normal vision.
Stimuli and procedure
We presented stimuli and collected responses on a Macintosh Mini computer using MATLAB and the Psychophysics Toolbox (Brainard, 1997); the displays appeared on a 17-in. CRT screen. Observers sat 60 cm from the screen.
The stimuli were six colored shapes presented equally spaced around the circumference of an imaginary circle centered at fixation with a radius of 4.2º. The fixation was a small white circle. Each item measured approximately 2.5º square. Each shape contained a white line, which was randomly oriented either vertically or horizontally. Each line measured 0.7º × 1º. There was one target (circle) and five distractors (triangle, square, and diamond) in each display. The identity of each distractor item was chosen randomly. The target as well as the distractors were green when the color singleton distractor was absent from the display; when a singleton distractor appeared, it could be red (RGB 255, 0, 0), yellow (RGB 255, 255, 0), purple (RGB 255, 0, 255), or orange (RGB 255, 150, 0).
The observers were to report the orientation of the line that appeared inside the green circle as quickly and accurately as possible. Observers responded “z” if the line was vertical and “m” if it was horizontal. Observers were informed that there might be a differently colored item present during some of the trials, but because the target was always green, they should ignore these items. Eye movements were not monitored, but observers were encouraged to maintain fixation. Each trial began with a fixation dot, visible for 1,000 ms. Next, the search display appeared and remained visible for 5,000 ms or until response. When observers failed to respond within 5,000 ms, they were encouraged to respond more quickly, and the trial was marked as incorrect. A beep was played following an incorrect response.
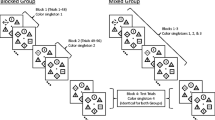
Observers first completed a 60-trial training block in which they searched for the green circle among heterogeneously shaped distractors in the absence of any color singletons. Following the training block, observers completed four test blocks of 48 trials each, as illustrated by Fig. 1. The test blocks were identical to the training block, except that one of the distractors was a color singleton on half of the trials. Each block had a differently colored color singleton. We counterbalanced the color singletons across the four block positions. Each block was preceded by a short, self-paced rest break.

Sequence of events for Experiment 1. Each search display was preceded by a 1,000-ms fixation point (not pictured) and then appeared on the screen for 5,000 ms or until response. Color singletons appeared in 50 % of the trials. The color of the color singletons changed between blocks of 48 trials
Results and discussion
RTs exceeding three standard deviations from an observer’s mean, incorrect RTs, and RTs following an incorrect response were removed from the analysis. This trimming eliminated less than 2 % of the data.
The results appear in Fig. 2. To evaluate whether a new color singleton initially slowed responses to the target, RTs were entered into a 2 × 2 repeated measures ANOVA, with Trial Order (RTs from the first or second half of a block) and Singleton Presence (present vs. absent) as factors. We found a main effect of trial order, F(1, 15) = 4.80, p < .05, with slower RTs in the first halves than in the second halves of blocks, and a marginally significant main effect of singleton presence, F(1, 15) = 3.62, p < .08, with slower RTs in singleton-present than singleton-absent trials. Most importantly, these main effects were subsumed by a two-way interaction between trial order and singleton presence, F(1, 15) = 21.72, p < .001. In planned comparisons, we found that RTs were longer in the presence of a singleton than in its absence during the first halves of blocks, t(15) = 5.26, p < .001, but not in the second halves of blocks, t(15) = 1.49, p > .16.

Response times (in milliseconds) as a function of trial order (first vs. second halves of blocks) and singleton presence (present vs. absent) across the four blocks of Experiment 1. The error rates of each condition are reported at the bases of the bars. Error bars represent 95 % within-subjects confidence intervals (Loftus & Masson, 1994)
To examine the generality of this pattern of results, we analyzed the amount of capture (RTs of singleton-present trials minus RTs of singleton-absent trials) in the first and second halves of each block. We performed a 2 × 4 repeated measures ANOVA with Trial Order (first or second half of a block) and Block (1, 2, 3, and 4) as the factors. There was no interaction between trial order and block, demonstrating that capture did not depend on whether observers were in the first or any other block, F(3, 45) < 1. Importantly, this test also indicates that capture did not diminish significantly over the experiment, as illustrated by Table 1. This analysis demonstrates that observers did not learn general distractor rejection in the first block, and even suggests that observers need experience with each individual color singleton in order to effectively suppress it.
Finally, we also thought it important to investigate more precisely the time course of this distractor tuning. To examine this, we conducted an even finer-grained microgenetic analysis by collapsing across all the blocks and dividing RTs into six sequentially ordered bins (i.e., Bin 1 was composed of the first four trials from each block, Bin 2 of the second set of four trials from each block, etc.). We submitted these bins to a 2 × 6 repeated measures ANOVA with Singleton Presence (present vs. absent) and Trial Order (Bins 1–6) as factors. The ANOVA found a marginally significant main effect of singleton presence, F(1, 15) = 3.62, p < .08. Mauchly’s test indicated that RTs under the trial order variable violated sphericity. Therefore, a Greenhouse–Geisser correction was applied to the p value of the analysis of trial order. There was no main effect of trial order, F(5, 75) = 1.89, p < .16. Importantly, both of these main effects were subsumed by the significant interaction between singleton presence and trial order, F(5, 75) = 5.97, p < .001. Post hoc analyses, using Tukey’s HSD critical value [Q .05(6, 75) = 4.16], revealed significant capture in the first bin, t(15) = 4.23, and the second bin, t(15) = 6.61, but no capture in the later bins, all ts < 1 (see Table 2). An informal examination of the individual trials also found this pattern of capture during the first eight to nine trials. This means that our division of the first and second halves of the blocks (bins of 12 trials) may have been conservative, as capture primarily occurred only during the first eight instances with a color singleton.
The mean error rate was 2.7 %, and we submitted arcsine-transformed error rates to the same 2 × 2 repeated measures ANOVA as the RTs. The only significant effect was of trial order, which revealed that observers were less accurate in the first than in the second halves of blocks, F(1, 15) = 5.84, p < .03, paralleling the RT results.
The present pattern of results strongly supports experience-dependent attentional tuning for distractor rejection. Although observers spent 60 trials searching for a well-specified target, which, presumably, engaged feature search mode, this experience with the target alone was insufficient to prevent capture by an irrelevant color singleton. Of course, one might argue that 60 trials were insufficient to fully enter feature search mode and exhibit distractor suppression. However, by the beginning of the fourth block, observers had encountered the target in the absence of a singleton distractor on 132 trials. Nevertheless, a new color singleton still captured attention and slowed RTs at the beginning of the fourth block, which argues against mere target exposure as being sufficient for distractor rejection.
Perhaps a more pressing concern with Experiment 1 is that the initial encounter with a new color singleton was perfectly confounded with the rest breaks. If attentional control was loosened or interfered with by a rest break, attention would be captured more readily at the beginning of a block than later in the block, and the present results might have nothing to do with encountering a new color singleton. Although past studies have reported persistence of attentional control settings across rest breaks, and even over weeklong delays (Leber & Egeth, 2006; Leber et al., 2009), in Experiment 2 we introduced new color singletons both following a break and midblock.
Experiment 2
In Experiment 2, we used a training block identical to that in Experiment 1, but we used two test blocks of 96 trials instead of four test blocks of 48 trials. In a single block of 96 trials, the first 48 trials of a block had one color singleton present on half of the trials, and the second set of 48 trials had a different color singleton present on half of the trials. Critically, the second color singleton was not preceded by a rest break. This design allowed us to compare capture by a new color singleton following a rest break to capture by a new color singleton that did not follow a rest break.
Method
Participants
A group of 16 University of Iowa undergraduates participated for course credit. All had normal or corrected-to-normal vision.
Stimuli and procedure
The stimuli and procedure were the same as in Experiment 1, except for the following changes. There was no error beep to alert observers of incorrect responses, and blocks now comprised 96 trials. This led to only two rest breaks in the experiment (one following the training block and one splitting the test blocks). Importantly, in the first quarter of each block, observers experienced a new color singleton after coming out of a rest break. During the third quarter of a block, observers experienced a new color singleton in midblock, not following a rest break.
Results and discussion
We treated RTs as in Experiment 1, and this trimming excluded less than 2 % of the data. The mean RTs appear in Fig. 3. We analyzed the data using a 4 × 2 within-subjects ANOVA, with Color Singleton Presence (present vs. absent) and Trial Order (first, second, third, or fourth quarter) as factors. There was a main effect of trial order, F(3, 45) = 4.28, p < .01, likely driven by longer RTs in the first than in the latter quarters of blocks. There was no overall main effect of singleton presence, F(1, 15) = 2.9, p > .10. Most importantly, there was an interaction between trial order and singleton presence, F(3, 45) = 2.75, p = .05, which paralleled the interaction in Experiment 1. Planned comparisons confirmed that observers were slower when the singleton was present than when it was absent during the first quarter of a block, t(15) = 2.39, p = .03, but not in the second quarter, t(15) < 1, n.s., consistent with Experiment 1. Planned comparisons also revealed that observers were slower during singleton-present trials than during singleton-absent trials in the third quarter of a block, t(15) = 2.47, p < .03, but not during the fourth quarter, t(15) < 1, n.s. These results from the last two quarters demonstrate that a new color singleton captured attention even when that color singleton was not preceded by a rest break. Furthermore, after sufficient experience with a color singleton (during the second and fourth quarters of blocks), distractors were effectively rejected and did not capture attention.

Response times (in milliseconds) as a function of trial order (first to fourth quarters) and singleton presence (present vs. absent). Note that the important comparison here is the presence of capture in both the first and third quarters of the blocks. The error rates of each condition are reported at the bases of the bars. Error bars represent 95 % within-subjects confidence intervals (Loftus & Masson, 1994)
We analyzed arcsine-transformed accuracy data using the same 4 × 2 repeated measures ANOVA as for the RTs. There was a marginally significant effect of trial order, F(3, 45) = 2.76, p = .054, but pairwise comparisons failed to find any significant differences in accuracy between quarters of the blocks, ps > .60. Other than this, the pattern of the accuracy data largely reflected that of the RT data, with lower accuracy when observers first experienced a color singleton.
The results of Experiment 2 indicated that the initial encounter with a color singleton produces capture, irrespective of that singleton’s position in a sequence of trials. Moreover, this capture occurred despite stimulus conditions that typically produce a highly selective attentional state in which capture, measured in aggregate RTs, is absent.
General discussion
These experiments investigated the underpinnings of effective distractor rejection. Although previous results had indicated that highly salient color singleton distractors could be rejected during a search task that required the target to be specified precisely (Bacon & Egeth, 1994), we found that a precise target template was not sufficient to prevent capture. Instead, observers were slower to discriminate a target in the face of a task-irrelevant color singleton—a hallmark signature of attentional capture—when the singleton first appeared during the course of the experiment. However, the distractor no longer slowed RTs after observers had a short amount of experience rejecting this distractor, thus replicating Bacon and Egeth’s results after sufficient experience with distractors. Importantly, we also found that capture persisted across blocks. Experience with distractor rejection does not appear to generalize to all color singletons, but is instead specific to the rejected item.
One point for discussion centers on previous results that have failed to find capture by a color singleton, even after a switch in the color of the color singleton (Leber et al., 2009). We believe that there are three possibilities for this lack of capture. First, these past experiments did not report their results as a function of block, which prevented a microgenetic analysis of capture as it unfolded across trials. Second, the previous work used an RSVP task to assess capture, and this task requires no visual search and involves no uncertainty about the target’s position. Third, both the target and distractors changed from block to block. This increased amount of change could have encouraged a more generalizable search strategy. These differences could allow for rapid tuning of distractor rejection, making it difficult to observe results similar to those we have reported here. These concerns notwithstanding, Leber has recently reexamined his results to explore whether capture occurred briefly after a color change, followed by efficient distractor rejection (i.e., feature search mode); his initial reanalyses showed data trends that appear to support our findings, with early capture and a rapid acquisition of distractor rejection (A. Leber, personal communication, November, 2011). However, these reanalyses did not find similar results across all of Leber et al.’s experiments. Furthermore, there are many differences between Leber et al.’s task and ours, and these differences might obscure the type of rapid distractor tuning that we have reported in the present experiments.
The present results bear a superficial similarity to a previous literature on novel pop-out. Repeated (i.e., familiar) words in four-word arrays produced poorer localization accuracy when appearing with unrepeated (novel) words, suggesting that the novel words popped out of the array and captured attention (Johnston, Hawley, Plewe, Elliott, & DeWitt, 1990). However, careful consideration of novel pop-out suggested that it could be explained without appealing to attentional capture (Christie & Klein, 1996). Other results have suggested an attentional preference for novel stimuli (Reicher, Snyder, & Richards, 1976), but these findings speak to familiarity in visual search generally, not to attentional capture by task-irrelevant distractors. One interesting development in the surprise capture literature is work investigating the role of expectancy in capture by novel color singletons (e.g., Horstmann, 2005). We believe our work to be related because the novel color singletons in our experiment may have captured attention because they violated expectancies, although we note that capture in our experiment extended beyond the first experience with the color singleton. This latter point indicates that novelty alone does not explain our results, because after the first encounter with a new singleton distractor, subsequent singletons are no longer novel.
Our data can also be applied to hypotheses in the attentional-capture literature. For example, Folk and Remington (1998) proposed that slowed RTs to the target when a color singleton is present in the additional-singleton paradigm reflect a filtering cost from the additional complexity of the display. It is possible that the slowed RTs to the target in the initial exposures to the color singleton in our experiment reflect a filtering cost and that this filtering cost diminishes with experience, but it is not clear why experience would diminish this filtering cost, given that the distractor is still present and would still need to be filtered. A second major account of singleton capture effects in the literature is Theeuwes’s (2010) disengage hypothesis. Under this hypothesis, attention is captured and must be disengaged from salient distractors before moving to the target; it is this disengagement that takes time and causes RTs to be slow in the presence of a color singleton distractor. It could be that observers in our experiment did not learn to suppress salient distractors, but instead became increasingly fast at disengaging from salient distractors. But, like the filtering hypothesis, it is not clear why disengagement would become faster with experience, unless one proposes that experience with the distractor itself affects attentional control and capture, which is precisely our view. Finally, there have been many demonstrations of priming in attentional capture (Lamy, Carmel, Egeth, & Leber, 2006; Olivers & Humphreys, 2003; Pinto, Olivers, & Theeuwes, 2005). We view our results as being largely consistent with this literature. For instance, our explanation that a lack of experience with a color singleton leads to capture is synonymous with the explanation that a color singleton not appearing in one of the preceding trials leads to capture.
Our results complement a growing literature demonstrating that experience broadly influences visual attention. Most relevant for the present experiments are results from Leber and colleagues (Leber & Egeth, 2006; Leber et al., 2009), who demonstrated that observers who are trained into feature search mode maintain that search mode even when the search displays change, the targets change, and the search can be performed as a simpler singleton search. These findings suggest that a well-specified, precise target template might be carried across an experiment and shape attentional capture. Our results suggest that a substantial component of so-called feature search mode involves learned distractor rejection. We hypothesize that the carryover of feature search mode in previous work (e.g., Leber et al., 2009) was due to distractor rejection. Had the singleton distractor changed color when the search displays changed, our experience-dependent distractor rejection account predicts that, initially, the learned feature search would have been insufficient to prevent capture by the new singleton. In fact, this prediction is supported by recent work from Zehetleitner, Goschy, and Müller (in press), who used a paradigm similar to that of Leber and Egeth (2006) and found that training with one color singleton in feature search mode did not prevent capture by a different color singleton in the transfer phase of the experiment.
Our present results fit well with the recently proposed signal suppression hypothesis of attentional capture (Sawaki & Luck, 2010, 2011). Under this account, salient distractors are detected, which generates an “attend to me” signal, but attention is prevented from visiting such salient items through active suppression. Evidence for this hypothesis comes from results showing the presence of a Pd event-related potential component in response to salient distractors. This component is thought to reflect distractor suppression (Hickey, Di Lollo, & McDonald, 2009), consistent with the idea that these distractors are detected but not attended (Sawaki & Luck, 2010). On the basis of our results, we suggest that signal suppression is the result of tuning via experience; distractors might initially produce an attentional shift, but the opportunity to suppress distractors bootstraps the suppression process, preventing capture. This scenario leads us to predict that early in an electrophysiological experiment, an N2pc index of capture would be present, followed by the gradual disappearance of this component with the simultaneous emergence of a Pd index of distractor suppression.
In sum, we interpret our results as suggesting that attentional control does not involve a simple dichotomy between stimulus-driven and goal-driven control. The role of experience must be taken into account, because goal-driven control can only be instantiated after sufficient experience with salient items in the display. Prior to this, stimulus-driven control might predominate.
References
Awh, E., Matsukura, M., & Serences, J. T. (2003). Top-down control over biased competition during covert spatial orienting. Journal of Experimental Psychology: Human Perception and Performance, 29, 52–63. doi:10.1037/0096-1523.29.1.52
Bacon, W. F., & Egeth, H. E. (1994). Overriding stimulus-driven attentional capture. Perception & Psychophysics, 55, 485–496. doi:10.3758/BF03205306
Brainard, D. H. (1997). The psychophysics toolbox. Spatial Vision, 10, 433–436. doi:10.1163/156856897X00357
Christie, J., & Klein, R. M. (1996). Assessing the evidence for novel popout. Journal of Experimental Psychology: General, 125, 201–207. doi:10.1037/0096-3445.125.2.201
Chun, M. M., & Jiang, Y. (1998). Contextual cueing: Implicit learning and memory of visual context guides spatial attention. Cognitive Psychology, 36, 28–71. doi:10.1006/cogp.1998.0681
Folk, C. L., & Remington, R. W. (1998). Selectivity in distraction by irrelevant featural singletons: Evidence for two forms of attentional capture. Journal of Experimental Psychology: Human Perception and Performance, 24, 847–858. doi:10.1037/0096-1523.24.3.847
Folk, C. L., Remington, R. W., & Johnston, J. C. (1992). Involuntary covert orienting is contingent on attentional control settings. Journal of Experimental Psychology: Human Perception and Performance, 18, 1030–1044. doi:10.1037/0096-1523.18.4.1030
Hickey, C., Di Lollo, V., & McDonald, J. J. (2009). Electrophysiological indices of target and distractor processing in visual search. Journal of Cognitive Neuroscience, 21, 760–775. doi:10.1162/jocn.2009.21039
Horstmann, G. (2005). Attentional capture by an unannounced color singleton depends on expectation discrepancy. Journal of Experimental Psychology: Human Perception and Performance, 31, 1039–1060. doi:10.1037/0096-1523.31.5.1039
Johnston, W. A., Hawley, K. J., Plewe, S. H., Elliott, J. M. G., & DeWitt, M. J. (1990). Attention capture by novel stimuli. Journal of Experimental Psychology: General, 119, 397–411. doi:10.1037/0096-3445.119.4.397
Kawahara, J. (2010). Identifying a “default” visual search mode with operant conditioning. Acta Psychologica, 135, 38–49. doi:10.1016/j.actpsy.2010.05.002
Kristjánsson, Á., & Campana, G. (2010). Where perception meets memory: A review of repetition priming in visual search tasks. Attention, Perception, & Psychophysics, 72, 5–18. doi:10.3758/APP.72.1.5
Lamy, D., Carmel, T., Egeth, H. E., & Leber, A. B. (2006). Effects of search mode and intertrial priming on singleton search. Perception & Psychophysics, 68, 919–932. doi:10.3758/BF03193355
Leber, A. B., & Egeth, H. E. (2006). It’s under control: Top-down search strategies can override attentional capture. Psychonomic Bulletin & Review, 13, 132–138. doi:10.3758/BF03193824
Leber, A. B., Kawahara, J., & Gabari, Y. (2009). Long-term abstract learning of attentional set. Journal of Experimental Psychology: Human Perception and Performance, 35, 1385–1397. doi:10.1037/a0016470
Loftus, G. R., & Masson, M. E. J. (1994). Using confidence intervals in within-subject designs. Psychonomic Bulletin & Review, 1, 476–490. doi:10.3758/BF03210951
Maljkovic, V., & Nakayama, K. (1994). Priming of pop-out: I. Role of features. Memory & Cognition, 22, 657–672. doi:10.3758/BF03209251
Olivers, C. N. L., & Humphreys, G. W. (2003). Attentional guidance by salient feature singletons depends on intertrial contingencies. Journal of Experimental Psychology: Human Perception and Performance, 29, 650–657. doi:10.1037/0096-1523.29.3.650
Pashler, H. (1988). Cross-dimensional interaction and texture segregation. Perception & Psychophysics, 43, 307–318. doi:10.3758/BF03208800
Pinto, Y., Olivers, C. N. L., & Theeuwes, J. (2005). Target uncertainty does not lead to more distraction by singletons: Intertrial priming does. Perception & Psychophysics, 67, 1354–1361. doi:10.3758/BF03193640
Reicher, G. M., Snyder, C. R., & Richards, J. T. (1976). Familiarity of background characters in visual scanning. Journal of Experimental Psychology: Human Perception and Performance, 2, 522–530. doi:10.1037/0096-1523.2.4.522
Sawaki, R., & Luck, S. J. (2010). Capture versus suppression of attention by salient singletons: Electrophysiological evidence for an automatic attend-to-me signal. Attention, Perception, & Psychophysics, 72, 1455–1470. doi:10.3758/APP.72.6.1455
Sawaki, R., & Luck, S. J. (2011). Active suppression of distractors that match the contents of visual working memory. Visual Cognition, 19, 956–972. doi:10.1080/13506285.2011.603709
Serences, J. T., Yantis, S., Culberson, A., & Awh, E. (2004). Preparatory activity in visual cortex indexes distractor suppression during covert spatial orienting. Journal of Neurophysiology, 92, 3538–3545. doi:10.1152/jn.00435.2004
Theeuwes, J. (1992). Perceptual selectivity for color and form. Perception & Psychophysics, 51, 599–606. doi:10.3758/BF03211656
Theeuwes, J. (2010). Top-down and bottom-up control of visual selection. Acta Psychologica, 135, 77–99. doi:10.1016/j.actpsy.2010.02.006
Zehetleitner, M., Goschy, H., & Müller, H. J. (in press). Top-down control of attention: It’s gradual, practice-dependent, and hierarchically organized. Journal of Experimental Psychology: Human Perception and Performance. doi:10.1037/a0027629
Author note
This research was supported in part by grants from the National Science Foundation (BCS 11-51209) and the National Institutes of Health (R01AG026027), and by research contracts from the Nissan Motor Corporation and the Toyota Motor Corporation.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Vatterott, D.B., Vecera, S.P. Experience-dependent attentional tuning of distractor rejection. Psychon Bull Rev 19, 871–878 (2012). https://doi.org/10.3758/s13423-012-0280-4
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13423-012-0280-4