
Abstract
Across many studies, researchers have found that representations in working memory (WM) can guide visual attention toward items that match the features of the WM contents. While some researchers have contended that this occurs involuntarily, others have suggested that the impact of WM contents on attention can be strategically controlled. Here, we varied the probability that WM items would coincide with either targets or distractors in a visual search task to examine (1) whether participants could intentionally enhance or inhibit the influence of WM items on attention and (2) whether cognitive control over WM biases would also affect access to the memory contents in a surprise recognition test. We found visual search to be faster when the WM item coincided with the search target, and this effect was enhanced when the memory item reliably predicted the location of the target. Conversely, visual search was slowed when the memory item coincided with a search distractor, and this effect was diminished, but not abolished, when the memory item was reliably associated with distractors. This strategic dampening of the influence of WM items on attention came at a price to memory, however, as participants were slowest to perform WM recognition tests on blocks in which the WM contents were consistently invalid. These results document that attentional capture by WM contents is partly, but not fully, malleable by top-down control, which appears to adjust the state of the WM contents to optimize search behavior. These data illustrate the role of cognitive control in modulating the strength of WM biases of selection, and they support a tight coupling between WM and attention.
Similar content being viewed by others

The influential biased-competition model of attention (Desimone & Duncan, 1995) posits that the active maintenance of an item in working memory (WM) results in top-down biasing of visual processing in favor of matching items over other, competing objects. This model explains how the holding of a search template will facilitate the selection of targets, but it also suggests that WM-matching stimuli might capture attention when they are not directly task relevant. For instance, if you are mentally rehearsing a reminder to buy milk on your way to the supermarket, you may find your attention caught by a milk advertisement, at the expense of attending to the road.
In many recent studies, researchers have found evidence for the guidance of attention by the contents of WM (see Soto, Hodsoll, Rotshtein, & Humphreys, 2008, and Olivers, Peters, Houtkamp, & Roelfsema, 2011, for reviews). These studies required participants to remember an item (such as a colored shape) while performing an intervening visual search task. The critical manipulation was whether the memory item reappeared in the search display, and if so, whether its location coincided with the search target or with an irrelevant distractor. In numerous studies (e.g., Downing, 2000; Olivers, Meijer, & Theeuwes, 2006; Soto, Heinke, Humphreys, & Blanco, 2005), it has been observed that participants are faster to complete a search when the WM item reappears at the location of the search target, and slower to find the target when the WM item reappears at the location of a distractor. This has led to the assertion that attention is captured by items that match the WM contents, even when those are irrelevant for the search task. Because this capture of attention has been observed in search for pop-out targets (Soto, Humphreys, & Heinke, 2006), and even when the memory items never predicted the search target location, some researchers have suggested that this may be an automatic effect (Olivers et al., 2006; Soto et al., 2005).
Other researchers have found no influence of memory-matching items (Downing & Dodds, 2004; Houtkamp & Roelfsema, 2006; Peters, Goeberl, & Roelfsema, 2008), and others have suggested that in certain conditions, participants can strategically avoid a distracting reappearing memory item (Han & Kim, 2009; Woodman & Luck, 2007). The WM capture effect has also been found to be eliminated in the presence of spatial precuing to the search target (Pan & Soto, 2010), as well as diminished by time constraints on the search task (Dalvit & Eimer, 2011) or by high WM processing loads (Soto & Humphreys, 2008). Thus, some controversy remains as to whether WM automatically biases the allocation of attention or whether top-down control can modulate the effect.
Carlisle and Woodman (2011) examined the automatic and strategic contributions to WM’s guidance of attention, following the logic of classic attention studies (Posner & Snyder, 1975). Participants were shown a colored memory cue and then completed a visual search among colored stimuli. After the search, they were shown a memory probe and asked whether it matched the memory cue. Participants experienced 20 %-, 50 %-, or 80 %-valid (i.e., when the WM-matching item corresponded with the search target) conditions. Carlisle and Woodman found that increased probabilities of valid trials amplified the costs of nonmatching targets, as well as the benefits of matching targets (although the benefits were more than twice as great), but several factors limit the implications of their findings for understanding the interplay between WM, attention, and cognitive control. As in most other studies, Carlisle and Woodman included a memory test after each search array. Thus, even when the WM item was likely to match a distractor, there might be motivation to attend to that item in order to refresh its representation (cf. Woodman & Luck, 2007). Furthermore, each of their conditions included some probability that the memory item would match the target, creating an incentive for orienting toward the item.
Here, we further examined the extent to which participants can flexibly use their knowledge about the validity of the memory contents to optimize search performance—namely, to boost attention toward WM-matching targets when the probability of validity is high, and to suppress the influence of the WM representation in the search process when the probability of invalidity is high. We varied the probability that search targets would occur at the locations of memory-matching stimuli, and we explicitly informed participants about these contingencies. Memory was only probed during surprise memory recognition trials, and never after the search task, so there was no incentive to attend to a reappearing memory item. Furthermore, we included a condition of 100 %-invalid memory cues, to examine whether WM items may still guide attention, even in the face of top-down effort to prevent their impact when they can only be harmful to the search. In a second experiment, we increased the WM difficulty to explore the give-and-take relationship between the memory and attention components of the task. For the first time, we examined how expectations about the relationship between memory items and search targets would in turn modulate the accessibility of WM representations. The study provides new insights for understanding how the reciprocal interaction between WM contents and attention may be modulated by cognitive control.
Experiment 1
Method
Participants
A group of 20 volunteers (11 female, 9 male) gave written informed consent and received course credit or a $10 payment for their participation. The study was approved by the Duke University institutional review board.
Experimental protocol
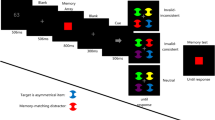
The experiment was run on a Dell Optiplex 960 computer using E-Prime (Version 2.0; Psychology Software Tools, 2007). The stimuli were viewed from approximately 60 cm on an LCD monitor with a 60-Hz refresh rate and a screen resolution of 1,280 × 1,024 pixels. Each trial began with the presentation of a white fixation dot at the center of the screen on a black background for 1,000 ms, followed by a blank screen for 500 ms, then a to-be-remembered colored circle cue, subtending approximately 1.3 ° in diameter, at the center of the screen for 250 ms (Fig. 1). A total of 2,000 ms after the offset of the cue, the target display appeared for 100 ms. The target display was composed of three colored circles—each subtending approximately 1.4 ° in diameter—at the corners of an imaginary triangle, with each corner approximately 2.5 ° from central fixation. Each circle contained a line of 0.57 ° length. Two of the lines were vertical and one—the target—was tilted 16 ° to the left or right. The participants’ task was to indicate the orientation of the line using a designated buttonpress to respond “left” or “right” tilted. Each target location and orientation occurred equally often and in a randomized order. To limit any incentive to attend to the memory item when it was irrelevant to the search task—while still ensuring that participants maintained the item in WM—20 % of the trials were catch trials on which, instead of a visual search display, the participants were given a recognition test for the memory item. The memory probes consisted of a colored circle 1.4 ° in diameter, presented at the center of the screen beneath a question mark for 1,500 ms. Participants had to indicate whether the probe was identical to the initial colored circle cue by using a designated buttonpress to respond “same” or “different.” Match and nonmatch probes occurred equally often, and their order was randomized. The memory items and search array circles were randomly selected from one of four colors, with RGB values as follows: red (155, 0, 0), blue (0, 0, 155), green (0, 165, 35), or yellow (170, 170, 45). Each color occurred equally often as a memory cue, and only one circle of each color appeared in the search display. Individual trials could be valid (the memory item reappeared surrounding the search target), invalid (the memory item reappeared surrounding a distractor), or neutral (the memory item did not reappear in the search display). The critical manipulation here, however, was that the trial conditions were blocked. Blocks could be composed of 100 % valid trials, 100 % invalid trials, 100 % neutral trials, or 50 % valid and 50 % invalid trials. The participants were given instructions at the beginning of each block informing them of these percentages.

Example trial sequence. The solid, dotted, and dashed lines represent different colors. Participants were asked to remember a colored circle over a delay, and then were either shown an array of three circles for a visual search or given a recognition test for the original cue. The memory test never occurred after the visual search display
The participants completed a practice session—comprising five trials of each condition, for a total of 20 trials—with a search duration of 500 ms, and then another practice session at the experimental search duration of 100 ms, to become acclimated to the speed of the search display. The participants then went on to complete four blocks of each condition, which occurred in random order. Each block consisted of 20 trials, for a total of 320 trials in all.
Results
The means and standard deviations for all measures are reported in Table 1. We examined response times (RTs) for correct visual search responses, according to block and trial condition (Fig. 2a). The mean performance on valid and invalid trials, in both the 100 %- and 50 %-predictable blocks, was normalized to the neutral baseline by subtracting the scores for each condition from the mean RT for the neutral blocks. These normalized RT scores were entered into a 2 × 2 ANOVA with the factors Validity (valid vs. invalid) and Predictability (100 % vs. 50 %). Search was faster overall on valid (vs. invalid) trials, F(1, 19) = 47.42, p < .001, η 2p = .71, and faster overall on 100 %-predictable blocks (vs. the less predictable 50 % validity) F(1, 19) = 16.97, p = .001, η 2p = .47. In other words, valid cues sped up the search process, and this speed-up was enhanced by cognitive control (cue predictability), whereas invalid cues slowed down the search process, and this slow-down was attenuated by cognitive control (Fig. 2b). The relative benefits of predictability, however, were comparable for valid and invalid trials, F(1, 19) = 1.31, p = .267, η 2p = .064.

(a) Experiment 1 visual search response times (RTs, in milliseconds) as a function of block and trial condition. (b) RT scores on each condition subtracted from the neutral baseline. Positive values reflect RTs faster than neutral, and negative values reflect RTs slower than neutral. Error bars reflect the mean standard errors
Search trial accuracy was at ceiling (mean = 97 %) and displayed no main effect of validity (p = .30) or predictability (p = .81), nor an interaction (p = .69). Neither RTs nor accuracy for memory catch trials varied significantly according to block condition (all ps > .1), and accuracy on the memory probes was high (93.4 %), confirming that participants were indeed keeping the cues in WM.
Discussion
The results clearly indicated both voluntary and involuntary contributions to the capture of attention by WM contents. While search speed was always fastest on validly cued trials, it was further augmented in the context of 100 %-valid blocks, when it was known that the WM item would coincide with the search target, and could be strategically enhanced. Conversely, search was always slowest on invalidly cued trials, but the impact of a memory-matching distractor was dampened in 100 %-invalid blocks, when it was known to be obstructive, and could be intentionally suppressed. However, 100 %-invalid blocks were still slower than neutral blocks, t(19) = 2.17, p < .05, indicating that participants could not completely prevent interference from the memory item. Unlike Carlisle and Woodman (2011), we found the benefits of valid trials and the costs of invalid trials to be of equal magnitudes and to be equivalently modulated by greater predictability. Because we included a 100 % antipredictive condition and precluded a memory-refreshing account by testing memory only on catch trials, our participants may have been able to more effectively dampen the impact of invalid cues—as opposed to the 80 % condition used by Carlisle and Woodman, in which participants might still have had some incentive to attend to invalid distractors to aid in the subsequent memory test.
We note that our 50 %-valid/50 %-invalid condition was somewhat predictive relative to a 33 %-valid condition, which would conform more to an “unpredictable” context, given our search set size of three. Previous research has indicated, however, that the effect of invalidity (invalid RT – neutral RT) should not necessarily vary with such a difference in the probability of validity. For instance, Carlisle and Woodman (2011, Exp. 2) showed comparable costs of invalidity, regardless of whether a block contained 20 % or 80 % valid trials. Likewise, Soto et al. (2005) observed no difference in the magnitude of invalidity costs, regardless of whether trials were 33 % valid/ 33 % invalid/33 % neutral or only invalid and neutral. The critical difference that we observed was between a condition that had some probability of validity (50 % valid/50 % invalid) and one with no valid trials (100 % invalid). The slight predictiveness of 50 % validity should not undermine the relevance of our observation—namely, that the interference effect from invalid WM cues was diminished, but not fully abolished, in the context of fully predictable invalid cues that should have maximized the application of cognitive control. We next asked whether cognitive control can modulate the state of the mnemonic contents in addition to the strength of the WM bias.
Experiment 2
Experiment 1 was not designed to investigate WM performance. Memory was tested on only 20 % of the trials, and the task was sufficiently easy that accuracy was very high (93 %). Neither accuracy nor RTs for the memory probes varied significantly with block condition. In Experiment 2, we sought simply to make the WM component more challenging—by expanding the color space from which stimuli could be drawn—so that accuracy would not be at ceiling, and so we might therefore observe variations in memory performance as a function of the strategic modulation of WM biases that occurred in Experiment 1.
Most prior studies have been conducted from the perspective that WM influences the allocation of attention, and typically they only addressed performance on the search task, without scrutinizing memory performance (but see Grecucci, Soto, Rumiati, Humphreys, & Rotshtein, 2010; Woodman & Luck, 2007). Here, we sought to more thoroughly understand the relationship between the memory and attention components of the task by probing the role of blockwise cue predictiveness on the memory contents themselves. This allowed us to adjudicate between two possible interpretations of the strategic control effects observed in Experiment 1 and in previous studies. On the one hand, the role of cognitive control in reducing the impact of WM items on attentional allocation could be seen to demonstrate a decoupling of WM and attention, thus supporting the independence of these processes (Peters et al., 2008; Woodman & Luck, 2007). On the other hand, strategic control may instead reflect modulation of the status of the memory representation itself, which in turn would amplify or diminish its effect on visual search. From the latter perspective, any effect of cognitive control on WM biases of selection would not be an expression of the independence between WM and attention, but of their reciprocity.
In conducting Experiment 2, we reasoned that if the second interpretation is correct, the strategic modulation of the WM contents’ impact on visual search should manifest itself in variations in the speed of memory recognition performance. Such a finding would also be consistent with a recent proposal that has reconciled disparate findings on the relationship between WM contents and attentional selection. Olivers et al. (2011) theorized that WM items can be designated different statuses or activation states and that only actively held items will influence perception. Other items can be appointed to an accessory state that will not impact visual attention (or, at least, not as considerably) but can then be retrieved into the active state when they become task relevant. If cognitive control over the impact of WM contents modifies the activation state of the critical memory representations, we would expect that an attempt to suppress a potentially interfering WM item would result in its assignment as an accessory item. This would then require a more time-consuming reinstatement into active memory in the event of a surprise recognition test. Thus, we predicted slower, but not necessarily less accurate, recognition memory responses in the 100 %-invalid than in the 100 %-valid cuing condition.
Method
Participants
A group of 28 volunteers (14 female, 14 male) gave written informed consent and received course credit or a $10 payment for their participation. The study was approved by the Duke University institutional review board.
Experimental protocol
The trial sequence in Experiment 2 was identical to that in Experiment 1, except that the proportion of memory trials was increased from 20 % to 50 %, and the memory test was made more difficult by increasing the number of colors from which the stimuli could be drawn. After the memory cue, participants completed a visual search on half of the trials and were tested for their memory of the cue color on the other half. The order of these trial types was random. Experiment 1 had utilized stimuli of four distinct colors, while in Experiment 2 we used those same four hues, but with three shades of each hue, for a total of 12 colors, with RGB values as follows: three shades each of red (180, 0, 0; 130, 50, 50; 163, 17, 62), blue (7, 186, 249; 1, 104, 255; 7, 70, 249), green (0, 255, 0; 1, 155, 0; 0, 80, 0), and yellow (252, 243, 62; 209, 204, 0; 255, 187, 51). The colors were selected from a range of shades that were determined in informal testing to be subjectively equally distinguishable, and the occurrence of each color was counterbalanced across conditions. When memory was tested for the color of the cue stimulus, the probe shade was either an exact match of the memory color or a different shade of the same hue, thus necessitating a more fine-grained visual WM representation than the cues in Experiment 1. When a memory color reappeared in the search array, it was only ever an exact match, never a different shade of the same color.
The participants completed a practice session—comprising five trials of each condition, for a total of 20 trials—with a search display duration of 500 ms, and then another practice session at the experimental search display duration of 100 ms. The experiment then consisted of two blocks each of the 100 %-valid, 100 %-invalid, and 100 %-neutral conditions, and four blocks of the 50 %-valid/50 %-invalid condition, all of the blocks presented in a random order. Each block constituted 32 trials, for a total of 320 trials in all.
Results
The means and standard deviations for all measures are reported in Table 2. The memory difficulty manipulation was successful at decreasing accuracy on memory trials (while keeping it well above chance). Where the overall memory accuracy in Experiment 1 had been 93.4 %, in Experiment 2 the value was significantly lower, at 77.2 %, t(46) = 6.76, p < .001. Despite this dramatic dip in WM accuracy, the pattern of search trial RTs in Experiment 2 was similar to that in Experiment 1 (Fig. 3a). Again, the scores were normalized—by subtraction from the neutral baseline—and entered into a 2 × 2 ANOVA with the factors Validity (valid vs. invalid) and Predictability (100 % vs. 50 %). Again, search was faster overall on valid (vs. invalid) trials, F(1, 27) = 85.5, p < .001, η 2p = .76, and faster overall on 100 %-predictable (vs. 50 %-validity) blocks, F(1, 27) = 10.24, p < .001, η 2p = .28, replicating the main findings of Experiment 1: Valid cues sped up search, and this speed-up was enhanced by cognitive control (cue predictability), whereas invalid cues slowed down search, and the slow-down was attenuated by cognitive control. A Validity × Predictability interaction, F(1, 27) = 7.57, p < .05, η 2p = .22, however, also revealed that the effect of predictability was greater for valid trials (100 % valid – 50 % valid) than for invalid trials (100 % invalid – 50 % invalid), t(27) = 2.75, p < .05 (cf. Carlisle & Woodman, 2011).

(a) Experiment 2 visual search response times (RTs, in milliseconds) as a function of block and trial condition. (b) Visual search RTs on each condition subtracted from the neutral baseline. Positive values reflect RTs faster than neutral, and negative values reflect RTs slower than neutral. (C) Memory probe RT for each block condition, subtracted from the neutral baseline. Error bars reflect the mean standard errors
Our task design left open the possibility that the WM bias in the 100 %-invalid blocks could reflect carryover from having previously experienced blocks with valid trials. To refute this possibility, we conducted a further analysis on the 12 participants who experienced the 100 %-invalid condition as their first block. We conducted the same ANOVA as above and again found main effects of validity, F(1, 11) = 56.21, p < .001, η 2p = .84, and predictability, F(1, 11) = 5.34, p < .05, η 2p = .33, as well as an interaction between the two, F(1, 11) = 6.88, p < .05, η 2p = .39. A two-tailed t test confirmed that even these 12 participants were significantly slower on 100 %-invalid blocks than on neutral blocks, t(11) = 5.74, p < .001. Thus, the capture of attention by the contents of WM in the 100 %-invalid blocks cannot be explained by carryover effects.
Search accuracy was sensitive to the WM validity condition in Experiment 2. While the ANOVA revealed neither a main effect of predictability (p = .25) nor a Validity × Predictability interaction (p = .7), there was a main effect of validity, F(1, 27) = 12.05, p < .01, η 2p = .31, reflecting better overall search accuracy when the WM cue coincided with the search target, as opposed to a distractor.
Finally, our main goal for Experiment 2 was to examine whether WM performance, now exposed to higher demands, would be affected by the experimental manipulations. And indeed, memory probe RTs did fluctuate with block condition. In a repeated measures ANOVA with the three-level factor of Block ConditionFootnote 1 (100 % valid, 50 % valid/50 % invalid, and 100 % invalid), there was a main effect of condition, F(2, 54) = 3.42, p < .05, η 2p = .11, and a significant linear trend, F(1, 27) = 4.99, p < .05, η 2p = .16. In line with our prediction that memory responses would be faster in the 100 %-valid blocks than in the 100 %-invalid blocks, a two-tailed t test confirmed this to be the case, t(27) = 2.23, p < .05. Memory recognition for accurate trials was fastest in the 100 %-valid blocks, then came the 50 %-valid blocks, and recognition was slowest in the 100 %-invalid blocks (Fig. 3c). Memory accuracy, on the other hand, did not differ according to block condition (p = .32).
Discussion
Although the memory task was more difficult in Experiment 2, participants displayed the same general search RT pattern as Experiment 1, with one important difference: Trial validity and block predictability interacted. When the memory demands were greater, the impact of predictability was diminished for invalid trials. We propose that the requirement to maintain a more precise visual memory representation magnified this representation’s impact when it was valid, but somewhat prevented the strategic attenuation of its impact when it was invalid. This enhanced WM representation in Experiment 2 may have also interfered on invalid trials beyond just slowing down the response, to the point that it increased error rates.
Of more direct concern for our hypothesis, however, is the memory performance. While memory accuracy was not modulated by validity or predictability, the participants took longer to retrieve and report memory items when they were strategically attempting to inhibit these items’ impact on visual search. We found no significant difference, however, in memory RTs between the 100 %-valid and neutral blocks (p = .9). This may well reflect a ceiling effect in the speed with which participants could respond to the memory probe. We predicted that the memory cues in an invalid context should be relegated to a different state, leading to slower RTs relative to the valid cues, which should remain accessible. This framework, however, makes no strong prediction about what should occur in the neutral context.
General discussion
Experiment 1 confirmed that both purposeful and unintentional contributions influence the capture of attention by the contents of WM. As in many other studies (Downing, 2000; Olivers et al., 2006; Soto et al., 2005), search performance was delayed in the presence of WM-matching distractors and was expedited by WM-matching targets. The magnitude of these costs and benefits, however, was modulated by the probability that WM cues would coincide with targets versus distractors. Despite the confidence with which participants could predict—and presumably attempt to avoid—invalid cues in 100 %-invalid blocks, however, they were still significantly slowed by these cues. In line with previous studies, these results suggest that WM biases of selection are modifiable by cognitive control (Carlisle & Woodman, 2011; Han & Kim, 2009; Woodman & Luck, 2007). This top-down influence over WM biases of selection is limited, however, because invalid-cue costs are still incurred under conditions of 100 % antipredictive cues.
Experiment 2 further extended our understanding of the nature of the relationship between WM and attention. We showed that memory recognition speed was sensitive to the validity and predictability of the WM item for visual search, further supporting the reciprocity of attention and WM processes; WM not only influenced the allocation of attention, but the status of the memory itself was modulated on the basis of the current goals. This pattern of memory performance indicates a possible means by which strategic control over the WM–attention interaction may be implemented for optimal performance. Specifically, it appears that control can occur at the level of the memory representation to alter its subsequent impact, and that this may be accomplished through manipulation of the status of the WM trace.
As Olivers et al. (2011) recently suggested, WM items may be maintained in different states of activation, and that status may determine how extensively they influence the allocation of attention. This possibility was first suggested by Downing and Dodds (2004), who found that an irrelevant memory item did not interfere with search, and it was supported by Olivers and Eimer (2011), who found that the extent of memory guidance depended on the imminence of the memory test. The present findings suggest that predictably helpful memory cues will be strategically, actively maintained in the focus of attention in order to promote faster search for a matching target, and that they will also be immediately accessible if memory is probed. Predictably invalid memory items, on the other hand, will be shifted to an accessory status, so as not to impede search performance, and will therefore require more time to restore to an active state for a memory probe response. This is consistent with the proposal made by Oberauer (2002) that an internal focus of attention within WM can hold a single representation at a time, and that a separate store can simultaneously contain several items for direct access. We propose that shifting between the internal focus of attention and the zone of direct access may be driven by the relevance of the WM contents vis-à-vis intermittent task goals.
Two important implications stem from these results. First, both the visual search and memory performance profiles indicate that a give-and-take relationship exists between the attention and WM components of this dual-task paradigm. When task demands necessitate a richly maintained memory representation, there are consequences for visual search performance that may be harmful or helpful, depending on the relationship between the memory contents and the search target. Likewise, when cognitive control is exerted to limit the impact of irrelevant memory contents on search performance, there are consequences for memory recognition. Thus, WM and attention processes appear to be tightly connected and not easily dissociable. Second, this cognitive control over WM biases is implemented (at least partly) through modification of the memory representation itself, and specifically via context-dependent enhancement or attenuation of the representation’s impact by shifting its WM state.
Notes
When analyzing search RTs, we conducted a 2 × 2 (Validity × Predictability) ANOVA. Memory trials, however, could not be analyzed in this way. Memory was only tested during catch trials, so there were no valid or invalid memory trials within the 50 %-validity blocks. Rather, we could only look at memory performance in the context of predictable versus unpredictable blocks. Consequently, in the memory analysis, we have entered each block condition of interest as a level in the ANOVA.
References
Carlisle, N. B., & Woodman, G. F. (2011). Automatic and strategic effects in the guidance of attention by working memory representations. Acta Psychologica, 137, 217–225. doi:10.1016/j.actpsy.2010.06.012
Dalvit, S., & Eimer, M. (2011). Memory-driven attentional capture is modulated by temporal task demands. Visual Cognition, 19, 145–153.
Desimone & Duncan. (1995). Neural mechanisms of selective visual attention. Annual Review of Neuroscience, 18, 193–222. doi:10.1146/annurev.ne.18.030195.001205
Downing, P. E. (2000). Interactions between visual working memory and selective attention. Psychological Science, 11, 467–473. doi:10.1111/1467-9280.00290
Downing, P. E., & Dodds, C. M. (2004). Competition in visual working memory for control of search. Visual Cognition, 11, 689–703.
Grecucci, A., Soto, D., Rumiati, R. I., Humphreys, G. W., & Rotshtein, P. (2010). The interrelations between verbal working memory and visual selection of emotional faces. Journal of Cognitive Neuroscience, 22, 1189–1200. doi:10.1162/jocn.2009.21276
Han, S. W., & Kim, M. (2009). Do the contents of working memory capture attention? Yes, but cognitive control matters. Journal of Experimental Psychology. Human Perception and Performance, 35, 1292–1302.
Houtkamp, R., & Roelfsema, P. R. (2006). The effect of items in working memory on the deployment of attention and the eyes during visual search. Journal of Experimental Psychology. Human Perception and Performance, 32, 423–442.
Oberauer, K. (2002). Access to information in working memory: exploring the focus of attention. Journal of Experimental Psychology: Learning, Memory, and Cognition, 28, 411–421.
Olivers, C. N. L., & Eimer, M. (2011). On the difference between working memory and attentional set. Neuropsychologia, 49, 1553–1558. doi:10.1016/j.neuropsychologia.2010.11.033
Olivers, C. N. L., Meijer, F., & Theeuwes, J. (2006). Feature-based memory-driven attentional capture: visual working memory content affects visual attention. Journal of Experimental Psychology. Human Perception and Performance, 32, 1243–1265.
Olivers, C. N. L., Peters, J., Houtkamp, R., & Roelfsema, P. R. (2011). Different states in visual working memory: when it guides attention and when it does not. Trends in Cognitive Sciences, 15, 327–333.
Pan, Y., & Soto, D. (2010). The modulation of perceptual selection by working memory is dependent on the focus of spatial attention. Vision Research, 50, 1437–1444.
Peters, J. C., Goeberl, R., & Roelfsema, P. R. (2008). Remembered but unused: the accessory items in working memory that do not guide attention. Journal of Cognitive Neuroscience, 21, 1081–1091.
Posner, M. I., & Snyder, C. R. (1975). Facilitation and inhibition in the processing of signals. In P. M. A. Rabbitt & S. Dornic (Eds.), Attention and performance V (pp. 669–682). New York, NY: Academic Press.
Psychology Software Tools, Inc. (2007). E-Prime (Version 2.0) [Software]. Sharpsburg, PA: Author.
Soto, D., Heinke, D., Humphreys, G. W., & Blanco, M. J. (2005). Early, involuntary top-down guidance of attention from working memory. Journal of Experimental Psychology. Human Perception and Performance, 31, 248–261.
Soto, D., Hodsoll, J., Rotshtein, P., & Humphreys, G. W. (2008). Automatic guidance of attention from working memory. Trends in Cognitive Sciences, 12, 342–348. doi:10.1016/j.tics.2008.05.007
Soto, D., & Humphreys, G. W. (2008). Stressing the mind: The effect of verbal suppression and cognitive load on attentional guidance from working memory. Perception & Psychophysics, 70, 924–934.
Soto, D., Humphreys, G. W., & Heinke, D. (2006). Working memory can guide pop-out search. Vision Research, 46, 1010–1018.
Woodman, G. F., & Luck, S. J. (2007). Do the contents of visual working memory automatically influence attentional selection during visual search? Journal of Experimental Psychology. Human Perception and Performance, 33, 363–377. doi:10.1037/0096-1523.33.2.363
Author note
This research was funded by National Institute of Mental Health Award R01MH087610 to T.E. and by a grant from the Medical Research Council (U.K., 89631) to D.S. We thank Alex Irwin for help with data collection.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Kiyonaga, A., Egner, T. & Soto, D. Cognitive control over working memory biases of selection. Psychon Bull Rev 19, 639–646 (2012). https://doi.org/10.3758/s13423-012-0253-7
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13423-012-0253-7