
Abstract
Emotion exerts varied influences on memory. While task-relevant item memory is often enhanced by emotion, associative memory is generally impaired. Unitization is known to improve associative memory, but its effects and mechanisms in protecting associative memory from emotional interference are rather obscure. The current study investigated associative memory by employing experimental manipulation of unitization (vs. nonunitization) encoding strategy and stimulus emotion (neutral, intrinsic negative, and extrinsic negative), combined with event-related potential (ERP) signatures of familiarity (FN400 old/new effects) and recollection (parietal late positive component/LPC old/new effects) in memory recognition. Both behavioral and ERP indices of associative recognition from the nonunitization group confirmed emotional interference in associative memory. Importantly, it was primarily intrinsic (vs. extrinsic) emotion that impeded associative memory. Unitization encoding improved memory performance in general, accompanied by enhanced recollection process and induction of familiarity process, which is typically not involved in associative memory recognition and was indeed absent in the nonunitization group. Importantly, unitization helped to preserve behavioral performance (specifically, response speed though not recognition strength) from interference by intrinsic emotion while largely reversed the detriment of intrinsic emotion on ERP indices of familiarity and recollection processes. Interestingly, a synergy between intrinsic emotion and unitization encoding was observed, which could underpin the facilitation of familiarity process in associative recognition of emotional pairs. Overall, current findings highlight interference by intrinsic emotion in associative memory, which is nonetheless responsive to mitigation by unitization encoding.
Similar content being viewed by others


Introduction
The notion that emotion influences memory is beyond dispute, but the ways in which this influence operates and manifests are varied and complex, remaining an area of hot debate in memory research (Easterbrook, 1959; Kensinger, 2009; Mackay et al., 2004; Mather & Sutherland, 2011). Emotion is known to influence multiple cognitive operations in diverse ways, generally facilitating intrinsic, task-relevant, and disrupting extrinsic, task-irrelevant ones (Pessoa, 2009; Williams, Mathews, & Macleod, 1996). Memory recruits various dissociable parallel processes, such as sensory/perceptual representation and contextual association, akin to the dual representation model of memory (Brewin, Gregory, Lipton, & Burgess, 2010). Therefore, depending on the inherent association (or the lack thereof) of these processes with emotion, memory is susceptible to both constructive and destructive forces of emotion (Bisby, Horner, Hørlyck, & Burgess, 2016).
Interestingly, in the realm of memory, while intrinsic emotion often facilitates item memory and feature binding of emotional items (Mather & Nesmith, 2008; Nashiro & Mather, 2011; Schmidt, Patnaik, & Kensinger, 2011; Steinmetz, Knight, & Kensinger, 2016), it tends to impair associative memory of discrete items by impeding association between individual emotional items (Madan, Caplan, Lau, & Fujiwara, 2012; Mao, You, Li, & Guo, 2015; Mather & Knight, 2008; Pierce & Kensinger, 2011; Rimmele, Davachi, Petrov, Dougal, & Phelps, 2011). In keeping with that, recent neuroimaging studies indicate that object perceptual processing in the sensory cortex is enhanced by emotion whereas associative binding in the hippocampus and parahippocampal gyrus is impaired by emotion (Luck, Leclerc, & Lepage, 2014; Mather et al., 2006; Murray & Kensinger, 2014; Onoda, Okamoto, & Yamawaki, 2009). To add to the complexity of emotional influence in memory, emotional effects on item memory per se could further differ drastically for central versus peripheral items (Kensinger, 2009; Mather, 2007), as the former being intrinsically, whereas the latter only extrinsically related with emotion. It is however not clear how intrinsic emotion compares with extrinsic emotion in influencing associative memory.
Nevertheless, emotional interference of associative memory has been shown to disappear or even turn into mnemonic benefits when associative processes are strengthened through effortful between-item integration during encoding (Guillet & Arndt, 2009; Murray & Kensinger, 2012). Unitization, an encoding strategy that integrates discrete items into a single unit, has been shown to enhance associative memory (Diana, Yonelinas, & Ranganath, 2008; Graf & Schacter, 1985, 1989). It makes intuitive sense that by strengthening associative binding during encoding, unitization can offset memory interference caused by emotion, thereby preserving associative memory. It also is possible that given the inherent emotional relation, discrete objects sharing the same emotion (e.g., two negative words) may receive privileged integration by unitization, affording a synergy between emotion and unitization to improve associative memory of emotional objects (Murray & Kensinger, 2014). That is, compared with extrinsic emotion, intrinsic emotion could exert a paradoxical facilitation effect on associative memory when unitization encoding is applied.
Familiarity and recollection represent two primary processes underlying memory recognition (Yonelinas, 2002), and the recollection process is known to play a major role in associative memory, particularly fostering recognition of relational bindings among items (Donaldson & Rugg, 1998; Hockley & Consoli, 1999; Opitz & Cornell, 2006; Yonelinas, 1997; Yonelinas, 2002). Nevertheless, when the unitization strategy is applied during encoding of between-item associations, familiarity can be engaged to support the recognition of the association as a unitized representation (Opitz & Cornell, 2006; Parks & Yonelinas, 2015). In keeping with that, event-related-potential (ERP) studies of associative recognition demonstrate that familiarity-based frontal old/new effect arises in the unitization (but not the nonunitization) condition (Tibon, Ben-Zvi, & Levy, 2014; Tibon, Gronau, Scheuplein, Mecklinger, & Levy, 2014; Tibon & Levy, 2014; Zheng, Li, Xiao, Broster, Jiang, et al., 2015). Therefore, insights into familiarity and recollection processes in emotion-laden associative memory would shed light on modulatory mechanisms, such as unitization, that can mitigate such emotional interference.
Therefore, we conducted an associative memory study using word pairs, where the encoding strategy was manipulated to either promote or prevent unitization. Specifically, to elucidate the mechanism underlying the interaction between emotion and unitization in associative memory, we assayed their impacts on the familiarity and recollection processes during the recognition of word pair associations, using their signature event-related potentials (ERPs). That is, the frontal old/new effect at 300~500 ms poststimulus, also known as the FN400 effect, was used to index familiarity-based recognition, while the parietal old/new effect in a late positive component (LPC; 500~800 ms poststimulus) was used to index recollection-based recognition (Curran, 2000; Diana, Yonelinas, & Ranganath, 2007; Rugg & Curran, 2007). Finally, as mentioned above, whether emotion is intrinsic or extrinsic may modulate how emotion and unitization influence associative memory. Stimulus emotional content was thus manipulated as intrinsic (involved in word pairs serving as the target) or extrinsic (involved in scenes serving as the context) to explicate emotional interference along the intrinsic-versus-extrinsic dimension.
Methods
Participants
Forty-eight college students (32 females; mean age 23.1 ± 2.2 years) from Capital Normal University participated in the experiment and received monetary compensation. Based on the typical size of similar effects in our lab (n = 17/group; f = 0.35; moderate to large; Huang, Yang, Zhang, & Guo, 2014; Liu, Rosburg, Gao, Weber, & Guo, 2017; Mao et al., 2015) and our design of three repeated measures and two independent groups, this sample size (n = 24/group) would have a reasonable power of 0.83. Participants were healthy, right-handed native Chinese speakers, with normal or corrected-to-normal vision. All of them were prescreened for history of neurological or psychiatric disorders, learning disorders, head injury or psychotropic drug use. Informed consent was obtained from each participant, which was approved by the Capital Normal University Institutional Review Board. Ten participants were excluded from the analyses, including three participants with very poor performance of the task (associative Pr < 0; detailed below), one with extremely low imagery success rate (<20% of pairs imagined), and six additional participants with insufficient valid ERP trials (n ≤ 15 in one or more of the experimental conditions). The final sample consisted of 38 participants, including 19 in the nonunitization group (14 females; mean age 24.2 ± 1.9 years) and 19 in the unitization group (14 females; mean age 23.2 ± 2.2 years).
Stimuli
Stimuli consisted of 320 pictures and 960 Chinese two-character words. Pictures served as the context for the word pairs at study, which depicted scenes with people, animals, landscapes, and artifacts. Pictures were selected from the International Affective Pictures System (Lang, Bradley, & Cuthbert, 2008), including 160 negative [valence: mean (SD) = 2.80 (0.71); arousal: mean (SD) = 5.84 (0.66), based on IAPS norms on a scale of 1~9] and 160 neutral [valence: mean (SD) = 5.20 (0.57); arousal: mean (SD) = 3.33 (0.65)]. Negative pictures were significantly more negative and arousing than neutral pictures [valence: t (159) = −33.41, p < 0.001; arousal: t (159) = 35.13, p < 0.001]. Pictures were matched for luminance and contrast by using adjustment curve in Adobe Photoshop 8.0. In addition, there was no difference between negative and neutral pictures (p’s > 0.05) in the objective measures of visual complexity (edge density and entropy; Marin & Leder, 2013).
For the word stimuli, we first selected 1,775 concrete nouns from the Modern Chinese frequency dictionary (Liu, 1990), with the mean frequency of 91.67 per million. An independent sample (N = 15) provided ratings of the words on a scale of 1 to 9 on the dimensions of valence, arousal, familiarity, and imageability. Based on the ratings, 960 words were selected, including 480 negative words [valence: mean (SD) = 3.21 (0.85); arousal: mean (SD) = 5.13 (1.30)] and 480 neutral words [valence: mean (SD) = 5.56 (0.54); arousal: mean (SD) = 3.07 (0.92)]. Negative words were more negative and arousing than neutral words [valence: t (479) = −48.94, p < 0.001; arousal: t (479) = 28.37, p < 0.001] but equal to neutral words on imageability, familiarity, and frequency (p’s > 0.05). Words were pseudo-randomly combined to form word pairs of negative/negative words or neutral/neutral words, resulting in 240 semantically unrelated negative and 240 semantically unrelated neutral word pairs.
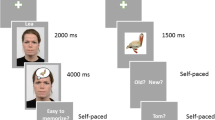
A total of 320 pairs (160 negative and 160 neutral pairs) were then combined with 320 pictures as materials for the study phase. The remaining 160 pairs (80 negative and 80 neutral pairs) served as new pairs during retrieval (Fig. 1A). The 320 picture-word-pair combinations formed 4 study lists of 80 each—“neg-neg”: a negative picture with a negative word pair, “neg-neu”: a negative picture with a neutral word pair, “neu-neg”: a neutral picture with a negative word pair, and “neu-neu”: a neutral picture with a neutral word pair. The “neg-neg” stimuli served as fillers, such that the word and picture stimuli could be fully counterbalanced across the groups. Given our interest in examining intrinsic versus extrinsic emotion (in relation to neutral emotion) in associative memory, this condition of mixed intrinsic and extrinsic emotion was not included in the analyses. At retrieval, 160 intact pairs (the same pairs at study), 160 rearranged pairs (by mixing words belonging to different study pairs), and 160 new pairs were presented, each containing 80 negative and 80 neutral pairs. The intact and rearranged pairs were randomly selected from each study list (half intact and half rearranged). The test pairs were counterbalanced across subjects to ensure that every word was presented equally often in intact or rearranged pairs. Careful examination on the picture-word-pair combinations and the rearranged pairs ensured minimal pre-existing semantic relatedness among the stimuli. As the old/new effect between intact and new pairs would be confounded by item memory, we focused on the comparison between intact and rearranged pairs, which would demonstrate a relatively pure old/new effect of associative memory (Li, Mao, Wang, & Guo, 2017; Rhodes & Donaldson, 2008; Zheng, Li, Xiao, Broster, Jiang, et al., 2015).

Example stimuli and experimental paradigm. (A) Example stimuli for each emotion category. Neutral pairs (target; e.g., “番茄 耳机” meaning “tomato-headphones”) were superimposed on negative image (context) to form the “neg-neu” condition. Negative pairs (e.g., “墓碑 鼻屎” meaning “tombstone-booger”) were superimposed on a neutral image as the “neu-neg” condition. Neutral pairs (e.g., “红旗 雨伞” meaning “flag-umbrella”) were superimposed on a neutral image as the “neu-neu” condition. The “neg-neg” condition (negative words on a negative image) only served as filled stimuli. (B) In the emotional associative memory task, participants completed a study phase followed by a test phase after a distraction task.
Procedure
Participants were seated at a distance of 60 cm from a Dell monitor in an electrically shielded room. Stimuli were displayed (using Presentation) at the center of the monitor against a gray background. The pictures subtended a visual area of 12° × 9°, and the word pairs presented in a 40-point black font subtended a visual area of 6.2° × 1.4°. A standard study-test paradigm was adopted, which included 8 study-test blocks, with each study phase followed by a test phase after a distraction task of 3-digit backward counting. Stimuli were thus split evenly into the 8 blocks, presented pseudo-randomly with no more than three consecutive trials from the same condition.
At study, each trial started with a black fixation cross for 1,000~1,500 ms, followed by a picture for 1,500 ms, during which the participants must verbally report whether they would “Back Away” or “Stay There” from the scene as they encountered them (Fig. 1B). A word pair was then superimposed at the center of the picture for 4,000 ms. During this period, participants were required to create a unitized (unitization group) or nonunitized (nonunitization group) mental imagery for the word pair as vividly as possible. Unitization involved creating an image of the words in a pair interacting with each other (e.g., for “office-eagle”: participants were asked to imagine some eagles in an office; Murray & Kensinger, 2012), whereas nonunitization involved creating a separate image for each individual word (e.g., for “office-eagle”: participants were asked to form an image of an office and to separately form an image of an eagle). Then, a screen with a prompt of “Please rate your success” was presented for 1,500 ms, and participants were to rate their success at generating mental images by button pressing (absolute failure, fuzzy imagery, moderately detailed imagery, and clear and vivid imagery). An ANOVA (emotion × strategy) on the proportion of appropriate mental images (pairs that were given a rating of “moderately detailed imagery” or “clear and vivid imagery”) revealed no main or interactive effect of emotion and strategy [F’s < 1.20, p’s > 0.31]. Therefore, imagery did not differ among the conditions. All participants had first completed a practice phase, including ten word pairs, where they were given examples and practiced on the encoding strategy of unitized or nonunitized mental imagery according to their group assignment (Murray & Kensinger, 2012). They were asked to describe the images they formed by oral report. We then assessed whether they formed the images successfully and provided feedback. All participants were able to use the encoding strategy appropriately after the practice phase.
At test, each trial started with a fixation cross for 1,000~1,500 ms, followed by a word pair for 2,000 ms. Pictures were not presented at retrieval. Participants were asked to make a response of “intact,” “rearranged,” or “new” via keyboard as quickly and accurately as possible. Buttons were counterbalanced across participants. Half of participants (in per group) made responses of “old” and “rearranged” by pressing the key “F” and “G” with left hand, and of “new” by pressing the key “J” with right hand. The other half of participants responded “old” and “rearranged” by pressing the key “H” and “J” with right hand, and “new” by pressing the key “F” with left hand.
EEG recording and analysis
The EEG was recorded using a 64-channel Neuroscan system with a 500-Hz sampling rate with a 0.05~100-Hz bandpass filter. The electrode locations adhered to the extended international 10–20 system. Electrooculogram (EOG) was recorded by a pair of electrodes placed outside the outer canthi of each eye and one infraorbital to the left eye. The left mastoid was used as the reference site online, and EEG signals were re-referenced offline to the average of the left and right mastoid recordings. Impedance was less than 5 kΩ. EEG/EOG signals were filtered with a bandpass of 0.05~40 Hz. EEG data from the test phase were separated into 1,100-ms epochs, including 100 ms before stimulus onset for baseline correction. Epochs with amplitude exceeding ±75 μV were rejected. EOG blink artifacts were corrected using a linear regression estimate (Gratton, Coles, & Donchin, 1983). The minimal number of trials for each condition was set at 16, and 6 participants were excluded for failing to meet the minimal number of trials.
Based on previous ERP investigations on frontal old/new effects and parietal old/new effects (Li et al., 2017; Rugg & Curran, 2007; Speer & Curran, 2007; Wolk et al., 2006), and based on visual inspection of the grand average ERP waveforms, time windows of 300~500 ms and 500~800 ms were isolated to characterize the frontal and parietal old/new effects, respectively. Mean amplitudes for these windows were obtained from the frontal (collapsed across F3, Fz, and F4) and the parietal (P3, Pz, and P4) scalp locations (Bader, Mecklinger, Hoppstädter, & Meyer, 2010; Zheng, Li, Xiao, Ren, & He, 2016) for statistical analysis.
Statistical analysis
Behavioral data of interest included reaction times (RTs) and a discrimination measure of old/new effects, associative Pr, defined by subtracting false alarm rates for rearranged pairs from hit rates for old/intact pairs (Jäger, Mecklinger, & Kipp, 2006; Snodgrass & Corwin, 1988). RTs were extracted from correct trials only (i.e., hits and correct rejections). ERP data of interest were extracted from correct trials only (Donaldson & Rugg, 1998; Paller et al., 2003) and included the frontal mean amplitude at 300~500 ms and the parietal mean amplitude at 500~800 ms. We conducted repeated analyses of variance (ANOVAs; with Greenhouse-Geisser correction for non-sphericity) with a between-subjects factor of encoding strategy (unitization or nonunitization) and two within-subjects factors of response (intact or rearranged) and emotion (neg-neu, neu-neg, or neu-neu) on the RT and ERP data. For the associative Pr index that was already adjusted for old/new (i.e., intact/rearranged) difference, we performed an ANOVA of strategy and emotion. P values in t test were corrected for false discovery rate (FDR) at p < 0.05.
Results
Behavioral results
Associative Pr
An ANOVA (emotion × strategy) on the associative Pr revealed a main effect of encoding strategy [F (1, 36) = 54.75, p < 0.001, η2p = 0.60]: greater associative Pr for the unitization than non-unitization group [t (36) = 7.4, p < 0.001], and a main effect of emotion [F (1.80, 64.89) = 5.44, p < 0.01, η2p = 0.13]: lower associative Pr for the intrinsic emotion (neu-neg) than extrinsic emotion (neg-neu) and neutral (neu-neu) conditions [t’s (37) > 2.99, p’s < 0.01; Fig. 2A]. However, the interaction between emotion and strategy was not significant [F (1.80, 64.89) = 0.03, p = 0.96]. Therefore, associative Pr was generally improved by unitization encoding and impaired by intrinsic emotion. The means and SDs of Pr for the different conditions, along with RTs and accuracy, are presented in Table 1.

Behavioral results. (A) Strength of associative memory (Pr) was enhanced by the unitization strategy but weakened by intrinsic emotion. (B) Similarly, RTs were speeded up by unitization but slowed down by intrinsic emotion with one exception (unitization remedied the RT slowdown by intrinsic emotion in the rearranged condition). *P < 0.05; **P < 0.01; ***P < 0.001. Error bars = S. E. M.
Response times
An ANOVA (emotion × response × strategy) on response times (RTs) revealed main effects of encoding strategy [F (1, 36) = 7.24, p < 0.05, η2p = 0.17], emotion [F (1.98, 71.24) = 5.30, p < 0.01, η2p = 0.13], and response [F (1, 36) = 172.97, p < 0.001, η2p = 0.83]. RTs were significantly faster in the unitization (vs. nonunitization) group, significantly slower in the intrinsic emotion (neu-neg) than extrinsic emotion (neg-neu) and neutral (neu-neu) conditions (p’s < 0.05) and significantly faster in the old/intact (vs. rearranged) condition (see means and SDs in Table 1). There also was an emotion-by-response interaction [F (2, 71.99) = 3.18, p < 0.05, η2p = 0.08] and, importantly, a three-way emotion-by-response-by-strategy interaction [F (2, 71.99) = 3.44, p < 0.05, η2p = 0.09; Fig. 2B]. We further broke down this three-way interaction in two follow-up ANOVAs.
A follow-up ANOVA (emotion × strategy) for intact responses revealed a main effect of encoding strategy [F (1, 36) = 6.99, p < 0.05, η2p = 0.16]: faster RTs for the unitization than nonunitization group. There also was a main effect of emotion [F (1.93, 69.33) = 3.12, p = 0.05, η2p = 0.08]: slower RTs for the intrinsic emotion (neu-neg) condition than the neu-neu condition (p < 0.05), whereas the extrinsic emotion (neg-neu) condition did not differ from the other conditions (p’s > 0.20). There was no emotion-by-strategy interaction [F (1.93, 69.33) = 0.23, p = 0.79]. Therefore, for intact responses, intrinsic emotion had a detrimental effect on recognition speed for both encoding strategies.
A similar ANOVA for rearranged responses revealed a main effect of encoding strategy [F (1, 36) = 6.74, p < 0.05, η2p = 0.16]: faster RTs for the unitization (vs. nonunitization) group. There also was an emotion effect [F (1.86, 66.84) = 6.07, p < 0.01, η2p = 0.14]: slower RTs for the intrinsic emotion (neu-neg) than neu-neu (p < 0.01) and neg-neu conditions (p < 0.05). Importantly, there was an interaction between encoding strategy and emotion [F (1.86, 66.84) = 3.32, p < 0.05, η2p = 0.08]. Further follow-up analyses revealed a main effect of emotion [F (2, 36) = 6.66, p = 0.01, η2p = 0.27] in the nonunitization group: slower RTs for the intrinsic emotion (neu-neg) than neu-neu (p < 0.01 FDR) and neg-neu conditions (p < 0.05 FDR) in the nonunitization group. In the unitization group, emotion, either intrinsic or extrinsic, did not exert an effect on response speed (p = 0.14). Therefore, for rearranged (i.e., new) responses, we observed a protective effect of unitization encoding in memory recognition speed.
ERP results
Frontal 300-500-ms window
An ANOVA (emotion × response × strategy) revealed a main effect of emotion [F (1.74, 62.76) = 12.57, p < 0.001, η2p = 0.26]: more positive waveforms in intrinsic emotion (neu-neg) than neu-neu and neg-neu conditions [t’s (37) > 4.20, p’s < 0.001], and a three-way emotion-by-response-by-strategy interaction [F (1.94, 69.82) = 3.52, p < 0.05, η2p = 0.09; Figs. 3 and 4].

ERPs in the unitization group. (A) Unitization group grand average ERP waveforms for the intact response (black) and rearranged response (red) in three conditions at two scalp locations (F3, Fz, F4 collapsed as the frontal site; P3, Pz, P4 as the parietal site). Indexing familiarity-based recognition of associative memory, the frontal waveforms during 300-500 ms (tan areas) differed between intact and rearranged conditions in the neu-neg and neu-neu conditions (i.e., FN400 effects). Indexing recollection-based recognition, parietal waveforms during 500-800 ms (grey areas) were stronger for the intact than the rearranged condition. (B) Topographical maps of the old/new effects (intact minus rearranged) in each time window.

ERPs in the non-unitization group. (A) Nonunitization group grand average ERP waveforms for the intact response (black) and rearranged response (red) in three conditions at two scalp locations. Indexing the absence of familiarity-based recognition effects in the nonunitization group, frontal waveforms during 300-500 ms (tan areas) were not significantly different between the intact and rearranged conditions. Indexing recollection-based recognition, parietal waveforms during 500-800 ms (grey areas) were stronger for the intact than the rearranged condition (i.e., parietal LPC or old/new effect), but only in the neu-neu condition. (B) Topographical maps of the old/new effects (intact minus rearranged) in each time window.
A follow-up ANOVA (emotion × response) for the unitization group revealed a main effect of emotion [F (1.87, 33.61) = 8.34, p < 0.001, η2p = 0.32]: more positive waveforms in the intrinsic emotion (neu-neg) than neu-neu and neg-neu conditions [t’s (18) > 2.90, p’s < 0.01]. There also was a main effect of response [F (1, 18) = 9.00, p < 0.01, η2p = 0.33]: more positive waveforms in the intact than rearranged condition, supporting an FN400 old/new effect. An interaction between emotion and response [F (1.65, 29.78) = 2.58, p < 0.05, η2p = 0.13] and the follow-up paired t tests revealed an old/new effect (intact vs. rearranged) in the intrinsic emotion (neu-neg) condition [t (18) = 3.07, p < 0.05 FDR] and neu-neu condition [t (18) = 3.25, p < 0.05 FDR], although not in the neg-neu condition (p = 0.62). A similar follow-up ANOVA for the nonunitization group revealed a main effect of emotion [F (1.65, 29.69) = 5.03, p < 0.05, η2p = 0.22]: more positive waveforms in the intrinsic emotion (neu-neg) than neu-neu and neg-neu conditions [t’s (18) > 2.49, p’s < 0.05]. However, no response effect (i.e., old/new effect) or emotion-by-response interaction was observed (p’s > 0.26).
Overall, besides a general emotion effect, there was an FN400 old/new effect in the unitization group in both the neu-neu and neu-neg (but not neg-neu) conditions but no old/new effects in the nonunitization group.
Parietal 500-800-ms window
A similar ANOVA (emotion × response × strategy) revealed a main effect of emotion [F (1.95, 70.02) = 16.78, p < 0.001, η2p = 0.32]: more positive waveforms in the intrinsic emotion (neu-neg) than neu-neu and neg-neu conditions [t’s (37) > 3.51, p’s < 0.01] and more positive waveforms in the neu-neu than neg-neu condition [t (37) = 1.94, p = 0.06; Figs. 3 and 4]. There also was a main effect of response [F (1, 36) = 17.42, p < 0.001, η2p = 0.33]: more positive waveforms in the intact than rearranged response condition [t (37) = 4.23, p < 0.001], indicative of an old/new memory effect. There was no main effect of strategy [F (1, 36) = 0.33, p = 0.57].
An interaction between emotion and response also emerged [F (1.96, 70.43) = 4.75, p < 0.05, η2p = 0.12]. Separate ANOVAs (emotion × response) for the two groups confirmed strong emotion and response effects in both groups (F’s > 4.38, p’s < 0.05 FDR), and an emotion-by-response interaction in the non-unitization group alone [F (1.96, 35.21) = 4.55, p < 0.05, η2p = 0.20]. Follow-up t tests between intact and rearranged conditions at each emotion level for the nonunitization group showed a significant old/new effect in the neu-neu condition [t (18) = 3.88, p < 0.01 FDR] but not the intrinsic (neu-neg; p = 0.92) or extrinsic emotion (neg-neu) condition (p = 0.34).
Discussion
The present study was designed to elucidate the effects of (intrinsic vs. extrinsic) emotion and unitization encoding strategy on associative memory. Our behavioral measures of memory recognition, Pr and RTs, converged on an overall benefit of unitization in contrast to a general detriment of intrinsic emotion for associative memory. Nevertheless, an important exception stood out: when the unitization strategy was applied, intrinsic emotion failed to slow down recognition of rearranged pairs, highlighting a protective effect of unitization encoding against emotional interference in associative memory. ERP results provided important insights into these behavioral findings. Reflecting enhanced familiarity process by unitization, FN400 (frontal 300~500 ms) old/new effects emerged in the unitization but not the nonunitization group for both neutral and negative word pairs. Supporting strengthened recollection, especially for word pairs associated with intrinsic or extrinsic emotion, parietal LPC (500~800 ms) old/new effects appeared for all word pairs in the unitization group but only for the neutral pair in the nonunitization group. Notably, relative to intrinsic emotion, extrinsic emotion had limited effects on associative memory, which also could be mitigated by unitization encoding.
In keeping with the notion that associative recognition recruits recollection but not familiarity (Donaldson & Rugg, 1998; Hockley & Consoli, 1999; Yonelinas, 1997), our results for the neutral (control) word pairs in the nonunitization group revealed parietal LPC old/new effects (p < 0.01) but not FN400 old/new effects (p = 0.53), confirming the engagement of recollection (vs. familiarity) in associative memory. Also consistent with the idea that familiarity would be engaged in associative memory following unitization encoding (Opitz & Cornell, 2006; Parks & Yonelinas, 2015), the unitization group showed both parietal LPC and FN400 old/new effects in the control condition (p ≤ 0.01). The close alignment of these ERP indices with existing theories and evidence thus corroborates their validity as distinct neural correlates of familiarity and recollection (Curran, 2000; Diana et al., 2007; Rugg & Curran, 2007).
Emotion effects in the nonunitization group replicated previous studies of associative memory involving emotional information. We observed that intrinsic emotion (neu-neg) reduced associative Pr and prolonged RTs, relative to the neutral (neu-neu) condition, supporting the idea of emotion disrupting associative memory (Madan et al., 2012; Mao et al., 2015; Mather & Knight, 2008; Pierce & Kensinger, 2011; Rimmele et al., 2011). Accordingly, in contrast to the neutral condition, there was no LPC old/new effect in the intrinsic emotion condition (Brewin et al., 2010). Our inclusion of an extrinsic (neg-neu) emotion condition, in which emotion was present in the context (background scenes), revealed that extrinsic emotion did not affect Pr and RTs (relative to the neutral condition), accentuating the detrimental impact of intrinsic emotion on associative memory. We also note that in the nonunitization group, the LPC old/new effect in the extrinsic emotion was not significant. Nonetheless, unlike the intrinsic emotion condition, there was a trend of greater LPC for the old than rearranged word pair (p = 0.34) in the extrinsic emotion condition. Given that the LPC component was the smallest in the extrinsic emotion condition, we speculate that contextual emotion may have disrupted this component in general and thus masked the old/new effect. Overall, these behavioral and ERP findings are consistent with multiple accounts arguing that emotional effects are particularly salient when the emotional information is intrinsic or of high priority (Bisby et al., 2016; Brewin et al., 2010; Madan et al., 2012; Mather & Sutherland, 2011; Pessoa, 2009; Stein, Zwickel, Ritter, Kitzmantel, & Schneider, 2009), thereby particularly potent at disrupting the binding of discrete items (Brewin et al., 2010; Luck et al., 2014; Mather et al., 2006; Murray & Kensinger, 2014; Onoda et al., 2009).
The employment of unitization encoding in the unitization group substantially improved associative memory. It is posited that the unitization encoding strategy can elicit the familiarity process in associative recognition by integrating separate items into a singular unit such that a unified representation is retrieved and recognized (Graf & Schacter, 1989). Indeed, as mentioned above, FN400 old/new effects emerged in the unitization group (while absent in the nonunitization group), which were accompanied by greater associative memory (Pr and RTs) in the unitization than nonunitization group, replicating recent reports of unitization enhancing familiarity and boosting associative memory (Ahmad & Hockley, 2014; Diana, Boom, Yonelinas, & Ranganath, 2011; Tibon, Ben-Zvi, et al., 2014; Tibon, Gronau, et al., 2014; Tibon & Levy, 2014; Tu & Diana, 2016; Zheng, Li, Xiao, Broster, Jiang, et al., 2015).
Of particular relevance to our investigation, unitization reversed the detrimental effect of emotion on both ERP indices of familiarity and recollection processes. The remedial effect of unitization on recollection was fairly equivalent for both intrinsic and extrinsic emotion conditions. That is, the parietal LPC old/new effects were restored for both emotional conditions, suggestive of a powerful, general effect of unitization in enhancing the recollection process. However, the effect of unitization on familiarity appeared to be unique to intrinsic emotion. In the unitization group, the intrinsic emotion condition elicited reliable familiarity (FN400 old/new effects), comparable in strength to the neutral condition, which were completely disrupted in the nonunitization group. In contrast, familiarity remained absent in the extrinsic emotion condition despite the unitization strategy. That intrinsic emotion commands an advantage in unitization over extrinsic emotion represents a synergy between intrinsic emotion and unitization in inducing familiarity during associative recognition. That is, unitization may leverage the inherent emotional association between negative word pairs, yielding a particularly strong effect of integration that underpins familiarity (Murray & Kensinger, 2014). In contrast, the presence of emotion in the context instead of the word items (i.e., extrinsic emotion) may disrupt the unitization process by distracting attention from the words to the context. Therefore, the synergy between unitization and intrinsic emotion in boosting the familiarity process would represent a paradoxical force counteracting with its disruptive impact on associative memory that likely extends beyond familiarity and recollection (Kensinger, 2009, emotion review). These two opposing forces could result in multifaceted behavioral outcome: ultimately, the balance of this tug-of-war could manifest in forms of memory improvement, deterioration, no effect. Indeed, the current behavioral outcome suggests that emotional interference was fairly predominant, outweighing unitization-related enhancement in recognition accuracy and response speed to old pairs, but unitization trumped emotional interference in recognition speed to rearranged pairs. This relatively specific effect on reaction time as opposed to recognition strength implicates enhanced processing fluency associated with strengthened familiarity via unitization (Jacoby & Whitehouse, 1989).
Prior studies have demonstrated effects of unitization in alleviating associative memory impairment by age and amnesia (Bastin et al., 2013; Diana, Yonelinas, & Ranganath, 2010; Giovanello, Keane, & Verfaellie, 2006; Quamme, Yonelinas, & Norman, 2007; Zheng, Li, Xiao, Broster, & Jiang, 2015; Zheng et al., 2016). Our results expand the application of unitization to emotion-induced associative memory impairment. Moreover, our experimental manipulation of unitization encoding strategy emphasizes a causal effect of unitization in this process. Given the possible synergy between emotion and unitization, unitization could be especially effective in mitigating emotional interference in associative memory. As such, strategies involving unitization encoding can be useful not only in everyday living but also as clinical interventions or self-help techniques in treating memory problems. Conversely, the knowledge that unitization can mediate persistent negative associations may promote novel clinical interventions of emotional disorders. For instance, treatment efficacy for posttraumatic stress disorder, represented by overgeneralized threat responses including traumatic flashbacks (Brewin et al., 2010), would improve when traumatic associative memory (e.g., between an innocuous place with a car accident) is dismantled by dissolving pathological integration of discrete events.
References
Ahmad, F. N., & Hockley, W. E. (2014). The role of familiarity in associative recognition of unitized compound word pairs. The Quarterly Journal of Experimental Psychology, 67(12), 2301–2324.
Bader, R., Mecklinger, A., Hoppstädter, M., & Meyer, P. (2010). Recognition memory for one-trial-unitized word pairs: Evidence from event-related potentials. Neuroimage, 50(2), 772–781.
Bastin, C., Diana, R. A., Simon, J., Collette, F., Yonelinas, A. P., & Salmon, E. (2013). Associative memory in aging: The effect of unitization on source memory. Psychology & Aging, 28(1), 275–283.
Bisby, J. A., Horner, A. J., Hørlyck, L. D., & Burgess, N. (2016). Opposing effects of negative emotion on amygdalar and hippocampal memory for items and associations. Social Cognitive & Affective Neuroscience, 11(6), 981–990.
Brewin, C. R., Gregory, J. D., Lipton, M., & Burgess, N. (2010). Intrusive images in psychological disorders: Characteristics, neural mechanisms, and treatment implications. Psychological Review, 117(1), 210–232.
Curran, T. (2000). Brain potentials of recollection and familiarity. Memory & Cognition, 28(6), 923–938.
Diana, R., Yonelinas, A., & Ranganath, C. (2010). Medial temporal lobe activity during source retrieval reflects information type, not memory strength. Journal of Cognitive Neuroscience, 22(8), 1808–1818.
Diana, R. A., Boom, W. V. D., Yonelinas, A. P., & Ranganath, C. (2011). ERP correlates of source memory: Unitized source information increases familiarity-based retrieval. Brain Research, 1367, 278–286.
Diana, R. A., Yonelinas, A. P., & Ranganath, C. (2007). Imaging recollection and familiarity in the medial temporal lobe: A three-component model. Trends in Cognitive Sciences, 11(9), 379–386.
Diana, R. A., Yonelinas, A. P., & Ranganath, C. (2008). The effects of unitization on familiarity-based source memory: Testing a behavioral prediction derived from neuroimaging data. Journal of Experimental Psychology: Learning, Memory, & Cognition, 34(4), 730–740.
Donaldson, D. I., & Rugg, M. D. (1998). Recognition memory for new associations: Electrophysiological evidence for the role of recollection. Neuropsychologia, 36(5), 377–395.
Easterbrook, J. A. (1959). The effect of emotion on cue utilization and the organization of behavior. Psychological Review, 66(3), 183–201.
Giovanello, K. S., Keane, M. M., & Verfaellie, M. (2006). The contribution of familiarity to associative memory in amnesia. Neuropsychologia, 44(10), 1859–1865.
Graf, P., & Schacter, D. L. (1985). Implicit and explicit memory for new associations in normal and amnesic subjects. Journal of Experimental Psychology: Learning, Memory, & Cognition, 11(3), 501–518.
Graf, P., & Schacter, D. L. (1989). Unitization and grouping mediate dissociations in memory for new associations. Journal of Experimental Psychology: Learning, Memory, & Cognition, 15(5), 930–940.
Gratton, G., Coles, M. G. H., & Donchin, E. (1983). A new method for off-line removal of ocular artifact. Electroencephalography & Clinical Neurophysiology, 55(4), 468–484.
Guillet, R., & Arndt, J. (2009). Taboo words: The effect of emotion on memory for peripheral information. Memory & Cognition, 37(6), 866–879.
Hockley, W. E., & Consoli, A. (1999). Familiarity and recollection in item and associative recognition. Memory & Cognition, 27(4), 657–664.
Huang, X., Yang, J. C., Zhang, Q., & Guo, C. (2014). The time course of spoken word recognition in Mandarin Chinese: A unimodal ERP study. Neuropsychologia, 63, 165-174.
Jacoby, L. L., & Whitehouse, K. (1989). An illusion of memory: False recognition influenced by unconscious perception. Journal of Experimental Psychology General, 118(2), 126-135.
Jäger, T., Mecklinger, A., & Kipp, K. H. (2006). Intra- and inter-item associations doubly dissociate the electrophysiological correlates of familiarity and recollection. Neuron, 52(3), 535–545.
Kensinger, E. A. (2009). Remembering the details: Effects of emotion. Emotion Review, 1(2), 99–113.
Lang, P. J., Bradley, M. M., Cuthbert, B. N. (2008). International affective picture system (IAPS): affective ratings of pictures and instruction manual (Rep. No. A–8).
Li, B., Mao, X., Wang, Y., & Guo, C. (2017). Electrophysiological correlates of familiarity and recollection in associative recognition: Contributions of perceptual and conceptual processing to unitization. Frontiers in human neuroscience, 11, 125.
Liu, Y. (1990). Dictionary of modern Chinese words in common uses. Beijing: Yuhang Publishing House.
Liu, Y., Rosburg, T., Gao, C., Weber, C., & Guo, C. (2017). Differentiation of subsequent memory effects between retrieval practice and elaborative study. Biological Psychology, 127, 134-147.
Luck, D., Leclerc, M. E., & Lepage, M. (2014). The potentiation of associative memory by emotions: An event-related FMRI study. Advances in Neuroscience, 2014 964024.
Mackay, D. G., Shafto, M., Taylor, J. K., Marian, D. E., Abrams, L., & Dyer, J. R. (2004). Relations between emotion, memory, and attention: Evidence from taboo stroop, lexical decision, and immediate memory tasks. Mem Cognit, 32(3), 474–488.
Madan, C. R., Caplan, J. B., Lau, C. S. M., & Fujiwara, E. (2012). Emotional arousal does not enhance association-memory. Journal of Memory & Language, 66(4), 695–716.
Mao, X., You, Y., Li, W., & Guo, C. (2015). Emotion impairs extrinsic source memory—an ERP study. Biological Psychology, 110, 182–189.
Marin, M. M., & Leder, H. (2013). Examining complexity across domains: Relating subjective and objective measures of affective environmental scenes, paintings and music. PloS One, 8(8), e72412.
Mather, M. (2007). Emotional arousal and memory binding: An object-based framework. Perspectives on Psychological Science, 2(1), 33–52.
Mather, M., & Knight, M. (2008). The emotional harbinger effect: Poor context memory for cues that previously predicted something arousing. Emotion, 8(6), 850–860.
Mather, M., Mitchell, K. J., Raye, C. L., Novak, D. L., Greene, E. J., & Johnson, M. K. (2006). Emotional arousal can impair feature binding in working memory. Journal of Cognitive Neuroscience, 18(4), 614–625.
Mather, M., & Nesmith, K. (2008). Arousal-enhanced location memory for pictures. Journal of Memory & Language, 58(2), 449–464.
Mather, M., & Sutherland, M. R. (2011). Arousal-biased competition in perception and memory. Perspectives on Psychological Science, 6(2), 114–133.
Murray, B. D., & Kensinger, E. A. (2012). The effects of emotion and encoding strategy on associative memory. Memory & Cognition, 40(7), 1056–1069.
Murray, B. D., & Kensinger, E. A. (2014). The Route to an integrative associative memory is influenced by emotion. PloS One, 9(1), e82372.
Nashiro, K., & Mather, M. (2011). The effects of emotional arousal on memory binding in normal aging and alzheimer’s disease. Am J Psychol, 124(3), 301–312.
Onoda, K., Okamoto, Y., & Yamawaki, S. (2009). Neural correlates of associative memory: The effects of negative emotion. Neuroscience Research, 64(1), 50–55.
Opitz, B., & Cornell, S. (2006). Contribution of familiarity and recollection to associative recognition memory: Insights from event-related potentials. J Cogn Neurosci, 18(9), 1595–1605.
Paller, K. A., Ranganath, C., Gonsalves, B., LaBar, K. S., Parrish, T. B., Gitelman, D. R., … Reber, P. J. (2003). Neural Correlates of Person Recognition. Learning & Memory, 10(4), 253-260.
Parks, C. M., & Yonelinas, A. P. (2015). The importance of unitization for familiarity-based learning. Journal of Experimental Psychology: Learning, Memory, & Cognition, 41(3), 881–903.
Pessoa, L. (2009). How do emotion and motivation direct executive control? Trends in Cognitive Sciences, 13(4), 160–166.
Pierce, B. H., & Kensinger, E. A. (2011). Effects of emotion on associative recognition: Valence and retention interval matter. Emotion, 11(1), 139–144.
Quamme, J. R., Yonelinas, A. P., & Norman, K. A. (2007). Effect of unitization on associative recognition in amnesia. Hippocampus, 17(3), 192–200.
Rhodes, S. M., & Donaldson, D. I. (2008). Electrophysiological evidence for the effect of interactive imagery on episodic memory: Encouraging familiarity for non-unitized stimuli during associative recognition. Neuroimage., 39(2), 873–884.
Rimmele, U., Davachi, L., Petrov, R., Dougal, S., & Phelps, E. A. (2011). Emotion enhances the subjective feeling of remembering, despite lower accuracy for contextual details. Emotion, 11(3), 553–562.
Rugg, M. D., & Curran, T. (2007). Event-related potentials and recognition memory. Trends in cognitive sciences, 11(6), 251–257.
Schmidt, K., Patnaik, P., & Kensinger, E. A. (2011). Emotion's influence on memory for spatial and temporal context. Cognition and Emotion, 25(2), 229–243.
Snodgrass, J. G., & Corwin, J. (1988). Pragmatics of measuring recognition memory: Applications to dementia and amnesia. Journal of Experimental Psychology: General, 117(1), 34–50.
Speer, N. K., & Curran, T. (2007). ERP correlates of familiarity and recollection processes in visual associative recognition. Brain Research, 1174(1), 97–109.
Stein, T., Zwickel, J., Ritter, J., Kitzmantel, M., & Schneider, W. X. (2009). The effect of fearful faces on the attentional blink is task dependent. Psychonomic Bulletin & Review, 16(1), 104–109.
Steinmetz, K. R. M., Knight, A. G., & Kensinger, E. A. (2016). Neutral details associated with emotional events are encoded: Evidence from a cued recall paradigm. Cognition and Emotion, 30(7), 1352–1360.
Tibon, R., Ben-Zvi, S., & Levy, D. A. (2014). Associative recognition processes are modulated by modality relations. Journal of Cognitive Neuroscience, 26(8), 1785–1796.
Tibon, R., Gronau, N., Scheuplein, A. L., Mecklinger, A., & Levy, D. A. (2014). Associative recognition processes are modulated by the semantic unitizability of memoranda. Brain & Cognition, 92, 19–31.
Tibon, R., & Levy, D. A. (2014). The time course of episodic associative retrieval: Electrophysiological correlates of cued recall of unimodal and crossmodal pair-associate learning. Cognitive, Affective, & Behavioral Neuroscience, 14(1), 220–235.
Tu, H. W., & Diana, R. A. (2016). Two are not better than one: Combining unitization and relational encoding strategies. Journal of experimental psychology: Learning, memory, and cognition, 42(1), 114–126.
Williams, J. M. G., Mathews, A., & Macleod, C. (1996). The emotional stroop task and psychopathology. Psychological Bulletin, 120(1), 3–24.
Wolk, D. A., Schacter, D. L., Lygizos, M., Sen, N. M., Holcomb, P. J., Daffner, K. R., & Budson, A. E. (2006). ERP correlates of recognition memory: Effects of retention interval and false alarms. Brain Research, 1096(1), 148–162.
Yonelinas, A. P. (1997). Recognition memory ROCs for item and associative information: The contribution of recollection and familiarity. Memory & Cognition, 25(6), 747–763.
Yonelinas, A. P. (2002). The nature of recollection and familiarity: A review of 30 years of research. Journal of Memory & Language, 46(3), 441–517.
Zheng, Z., Li, J., Xiao, F., Broster, L. S., & Jiang, Y. (2015). Electrophysiological evidence for the effects of unitization on associative recognition memory in older adults. Neurobiology of Learning & Memory, 121, 59–71.
Zheng, Z., Li, J., Xiao, F., Broster, L. S., Jiang, Y., & Xi, M. (2015). The effects of unitization on the contribution of familiarity and recollection processes to associative recognition memory: Evidence from event-related potentials. International Journal of Psychophysiology, 95(3), 355–362.
Zheng, Z., Li, J., Xiao, F., Ren, W., & He, R. (2016). Unitization improves source memory in older adults: An event-related potential study. Neuropsychologia, 89, 232–244.
Acknowledgements
The present study was supported by the National Natural Science Foundation of China (31671127), Capacity Building for Sci-Tech Innovation – Fundamental Scientific Research Funds (025185305000/200). We thank Bingcan Li for helpful comments. We thank Xue Yang, Jing Lu and Xinyang Liu for research assistance. We also thank Xi Chen and Zhengcao Cao for helping selecting experimental materials.
Author information
Authors and Affiliations
Corresponding authors
Rights and permissions
About this article

Cite this article
Han, M., Mao, X., Kartvelishvili, N. et al. Unitization mitigates interference by intrinsic negative emotion in familiarity and recollection of associative memory: Electrophysiological evidence. Cogn Affect Behav Neurosci 18, 1259–1268 (2018). https://doi.org/10.3758/s13415-018-0636-y
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13415-018-0636-y