
Abstract
In the present study, we considered error-related brain activity in event-related potentials, to investigate the relationship between error monitoring—that is, the detection and evaluation of erroneous responses—and action effect monitoring—that is, monitoring of the sensory consequences of behavior. To this end, participants worked on a task-switching paradigm that consisted of a free-choice task, in which a puzzle piece had to be attached to an existing one (the prime task), and a subsequent color flanker task (the probe task). We examined whether unexpected action effects in the prime task would affect the subsequent error monitoring in the probe task. We found the neural correlates of error monitoring during the probe task, the error-related negativity as well as the error positivity, to be increased after unexpected action effects in the prime task. In contrast, the neural correlates of visual attention were decreased after unexpected action effects, in line with recent findings on an attenuation of sensory processing after errors. Our results demonstrate a direct link between monitoring processes in the two tasks. We propose that both error monitoring and action effect monitoring rely on a common generic monitoring system related to novelty detection or affective processing. Preactivating this system by means of unexpected action effects increases the sensitivity for detecting an error in the subsequent task.
Similar content being viewed by others


The ability to flexibly adapt to changing task demands requires a complex system of monitoring processes that constantly check for deviations between the current behavior and the intended goals. The output of these monitoring processes is used for adjustments to optimize goal-directed behavior. In recent years, different lines of research have focused on different types of monitoring. Research on error monitoring has addressed the question of how the brain is able to detect errors and adjust attention and behavior in a way so that further errors are avoided (Ullsperger, Fischer, Nigbur, & Endrass, 2014). In contrast, research on action effect monitoring has focused on how the (predicted) sensory consequences of behavior (so-called action effects) are processed and how violations of these predictions are detected (Band, van Steenbergen, Ridderinkhof, Falkenstein, & Hommel, 2009; Cardoso-Leite, Mamassian, Schütz-Bosbach, & Waszak, 2010; Wirth, Janczyk, and Kunde, 2018). In the present study, we aimed to investigate the relationship between error monitoring and action effect monitoring. Our approach was to examine whether manipulating action effect monitoring in one task generalizes to error monitoring in a subsequent task. Such a result would provide support for the idea that both types of monitoring rely on common processes, or even represent instances of a generic monitoring system.
Research on error monitoring has mainly used scalp electroencephalographic methods to reveal the neural basis of error detection. When humans commit errors in a speeded-choice task, a cascade of event-related potentials (ERPs) is elicited, reflecting monitoring processes that detect and evaluate the significance of the error. These processes are mirrored by two distinct neural correlates, the error-related negativity (Ne/ERN; Falkenstein, Hohnsbein, Hoormann, & Blanke, 1991; Gehring, Goss, Coles, Meyer, & Donchin, 1993) and the error positivity (Pe; Falkenstein et al., 1991; Overbeek, Nieuwenhuis, & Ridderinkhof, 2005). The Ne/ERN has a frontocentral distribution and peaks within 100 ms after the response. It is generally considered to be generated in the medial frontal cortex (MFC; Gehring et al., 1993; Holroyd, Dien, & Coles, 1998; van Veen & Carter, 2002) and to represent either a response conflict between an error and a corrective response tendency (Yeung, Botvinick, & Cohen, 2004) or a reward prediction error subserving reinforcement learning (Holroyd & Coles, 2002). Both accounts can explain why the Ne/ERN is typically larger when errors are less frequent or less expected (e.g., Gehring et al., 1993; Jessup, Busemeyer, & Brown, 2010; Oliveira, McDonald, & Goodman, 2007), but mainly the latter idea receives support from the finding that negative feedback about the correctness of a response elicits a frontocentral negativity that strongly resembles the Ne/ERN (Holroyd & Coles, 2002). In contrast, the Pe peaks around 300–500 ms after the response over parietal electrodes and is linked to processes of error awareness (Nieuwenhuis, Ridderinkhof, Blom, Band, & Kok, 2001; Overbeek et al., 2005). Recent evidence has suggested that the Pe reflects the accumulation of evidence that an error has occurred (M. Steinhauser & Yeung, 2010) and is a correlate of response confidence (Boldt & Yeung, 2015).
The role of action effects for performance has been described by ideomotor theory (Hommel, Müsseler, Aschersleben, & Prinz, 2001; James, 1890; Shin, Proctor, & Capaldi, 2010), which assumes that humans produce actions by anticipating the intended sensory consequences of these actions. For instance, a key press in an experiment might be produced by anticipating the proprioceptive or visual consequences of this key press (Janczyk, Durst, & Ulrich, 2017; Janczyk & Lerche, 2018; Pfister & Kunde, 2013; Wirth, Pfister, Brandes, & Kunde, 2016). If motor actions can be selected using codes for their sensory consequences, it can reasonably be assumed that actors monitor the ultimately produced effects as well, to determine whether the intended consequences have actually been realized or to establish new action effect associations. Such an action effect monitoring process was proposed by Welford as early as 1952. Recently, evidence for action effect monitoring has been provided using behavioral and neurophysiological methods.
Wirth, Janczyk, and Kunde (2018) required participants to produce left–right responses that were followed by either a spatially compatible or incompatible action effect. On the basis of the observation that incompatible action effects are harder than compatible action effects to process perceptually (Desantis, Roussel, & Waszak, 2014), it has been assumed that incompatible effects might invoke the proposed effect-monitoring process longer than compatible effects do. In line with this idea, the production of incompatible action effects increased the response time (RT) to an immediately following stimulus more than compatible effects did. This observation suggests that effect monitoring is a process that interferes with responding to another stimulus, with a duration depending on specific effect features such as the compatibility between an action and effect.
A subsequent study revealed similar influences by manipulating the expectedness of action effects (Wirth, Steinhauser, Janczyk, Steinhauser, & Kunde, 2018). For example, enhanced action effect monitoring was found for infrequent incompatible action effects when they occurred among frequent compatible action effects, via increased RTs to a subsequent classification task. These behavioral observations were mirrored by an increased amplitude of the frontocentral P3a following such infrequent action effects. Because this ERP component is sensitive to the (un)expectedness of an event (Polich, 2007; Polich & Criado, 2006), the results suggest that responses to the subsequent task were delayed due to an increased allocation of attention to the unexpected action effect (see also Waszak & Herwig, 2007). Such an involvement of medial–frontal brain structures in action effect processing, notably the presupplementary motor area, has been supported by a number of neuroimaging and brain stimulation studies (Elsner et al., 2002; Hughes & Waszak, 2011; Melcher, Weidema, Eenshuistra, Hommel, & Gruber, 2008; Ticini, Schütz-Bosbach, Weiss, Casile, & Waszak, 2012; for a review, see Waszak, Cardoso-Leite, & Hughes, 2012).
Of course, error monitoring and action effect monitoring differ in crucial aspects. Most importantly, unlike errors or performance feedback, action effects do not necessarily carry information about the correctness of a decision or future reward. For instance, a response can be detected as an (unexpected) error by error monitoring, but still produce the anticipated proprioceptive and visual action effects. However, the two types of monitoring also share crucial conceptual and empirical characteristics. Both error monitoring and action effect monitoring serve to detect action outcomes. Moreover, both errors and action effects have been shown to elicit or modulate frontocentral ERP components, and in both cases the activity becomes greater when errors or action effects are less frequent or less expected. Particularly, the latter observation raises the possibility that both forms of monitoring partially rely on more generic monitoring mechanisms that detect novel or unexpected events.
The idea that error monitoring involves the detection of novel and unexpected events has been discussed before (Oliveira et al., 2007; Wessel, Danielmeier, Morton, & Ullsperger, 2012; Wessel, Klein, Ott, & Ullsperger, 2014). Novel and unexpected stimuli have been shown to elicit a negative frontocentral deflection similar to the Ne/ERN (Folstein & Van Petten, 2008; Näätänen & Gaillard, 1983), and both errors and novel stimuli lead to slowed responses on the subsequent trial (Barcelo, Escera, Corral, & Periáñez, 2006; Danielmeier & Ullsperger, 2011; Iwanaga & Nittono, 2010; Rabbitt & Rodgers, 1977). On the basis of evidence from ERP and fMRI data, Wessel and colleagues (Wessel & Aron, 2017; Wessel et al., 2012) argued that the same neural circuits are involved in monitoring erroneous responses and novel events.
First evidence that error monitoring and action effect monitoring might share common mechanisms can be derived from a study that compared ERPs elicited by action effects and by external feedback about the correctness of a response (Band et al., 2009). Feedback indicating an error has previously been shown to elicit a feedback-related negativity (FRN) with a frontocentral scalp distribution equal to the Ne/ERN, but peaking 250–300 ms after the onset of the feedback stimulus (Holroyd & Coles, 2002; Miltner, Braun, & Coles, 1997). Band et al. reported a negativity after unpredicted but otherwise task-irrelevant action effects that strongly resembled the FRN in a condition with task-relevant feedback. They interpreted this action effect negativity as reflecting the same reward prediction error that has been linked to the FRN, and they argued that the monitoring of task-irrelevant action effects and feedback monitoring resort to the same structures in the MFC. Since feedback processing and error monitoring have been discussed as involving similar processes (Holroyd & Coles, 2002), this would suggest that error monitoring and action effect monitoring also might share common mechanisms. Similarly, Gentsch, Ullsperger, and Ullsperger (2009) compared actual performance errors with externally triggered errors that were introduced as occasional malfunctions of the keyboard. They found that errors and malfunctions elicited temporarily distinguishable ERP components—an Ne/ERN and an FRN, respectively—that nonetheless were attributable to the same neural source through an independent component analysis (see also M. Steinhauser & Kiesel, 2011).
The goal of the present study was to provide direct evidence for a relationship between error monitoring and action effect monitoring. As we described above, previous studies have inferred commonalities between monitoring mechanisms by comparing the spatial and temporal characteristics of the underlying neural correlates. A shortcoming of that approach is that these neural correlates are rather unspecific, because similar ERPs, and even similar brain structures, can be linked to very different functions. Here we adopted a novel approach by asking whether manipulating one type of to-be-monitored event would exert influences on monitoring of the other event. Specifically, our hypothesis was that preactivating the action effect monitoring system in one task should increase the sensitivity of error monitoring in an immediately following subsequent task. Such a result would provide evidence for the idea that both types of monitoring are instances of a generic monitoring system or, at least, rely on common processes such as monitoring for novel and unexpected events.
To test this hypothesis, we employed a task-switching scenario based on the design employed by Wirth and colleagues (Wirth, Janczyk, & Kunde, 2018; Wirth, Steinhauser, et al., 2018), in which a prime task and a probe task were performed in close succession. The prime task required participants to give a free-choice response (upper or lower key) that was followed by a visual action effect. This action effect was presented in either the direction corresponding to the response (compatible) or the opposite direction (incompatible). Crucially, we manipulated the expectedness of the action effect by mixing either frequent compatible and infrequent incompatible action effects, or vice versa, within blocks of trials. After a brief interval, the stimulus of the probe task (a flanker task) was presented, and participants had to classify this stimulus with an error-prone speeded response. We assumed that an unexpected action effect in the prime task would be accompanied by enhanced action effect monitoring, and that this in turn would preactivate error monitoring in the probe task. We therefore predicted that unexpected action effects in the prime task would lead to a larger Ne/ERN or Pe in the probe task. In addition, we investigated the effect of unexpected action effects in the prime task on sensory processing in the probe task. Recent research has indicated that performance errors attenuate attentional processing in subsequent tasks (Buzzell, Beatty, Paquette, Roberts, & McDonald, 2017; Van der Borght, Schevernels, Burle, & Notebaert, 2016). An equivalent finding on unexpected action effects would provide additional evidence for the account that both types of monitoring are part of one common monitoring system.
Method
Participants
Twenty-four female participants, between 18 and 26 years of age (mean = 22.1), took part in the experiment and received either monetary compensation (€8 per hour) or course credit. All participants reported normal or corrected-to-normal vision and hearing, had no history of neurological or psychiatric diseases, and were naïve concerning the hypotheses of the experiment. They provided written informed consent prior to the experiment. The experiment was conducted in accordance with the Declaration of Helsinki, and the study was approved by the ethics committee of the Catholic University of Eichstätt-Ingolstadt.
Tasks and procedure
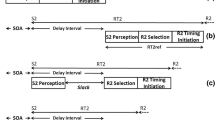
The paradigm was adapted from that of Wirth et al. (Wirth, Janczyk, & Kunde, 2018; Wirth, Steinhauser, et al., 2018). The stimuli for the prime task (S1) were bright gray pictures of puzzle pieces with connectors at the top and bottom, presented centrally on the screen at a visual angle of 4.7° and a viewing distance of 70 cm (Fig. 1). Participants were asked to add an equal-sized puzzle piece (the action effect E) at either the top or the bottom of S1 with a left-hand key press on either the upper (“A”) or lower (“Y”) key of a German QWERTZ keyboard.Footnote 1 The prime task was free choice, so participants could choose freely whether to press the top or the bottom key. Nonetheless, they were instructed to maintain an approximately equal ratio of top to bottom responses (see, e.g., Naefgen, Dambacher, & Janczyk, 2017). The subsequent probe task required participants to respond to target color squares (S2; red vs. blue vs. yellow; visual angle of 1.2° per square) presented centrally within the original puzzle piece (S1), with always incongruent flanker color squares; this target required a right-hand response on the “B,” “N,” or “M” keys (see also Fig. 1). The target was presented after the flankers in order to increase the difficulty, and thus the probability of errors, in the probe task. The letter–response mapping was counterbalanced between participants. All stimuli were presented against a black background.

Trial procedure of the experiments. In the prime task, a puzzle piece had to be added at the top or the bottom of a centrally presented piece by pressing an upper or lower button. This response started the response–stimulus interval (RSI), after which a second stimulus appeared within the puzzle piece (color flanker task) and had to be classified in the probe task.
The timing of a trial is illustrated in Fig. 1. Each trial started with a fixation cross presented randomly for 450 to 550 ms, followed by S1. Immediately after the response to the prime task, E was presented, and S1 and E remained on the screen until the end of the trial, to prevent offset effects in the ERPs. If after a maximum of 2,000 ms no key had been pressed, the trial counted as an omission, and no E was displayed. To reduce the likelihood of anticipatory responses in Task 2, a response–stimulus interval (RSI) of 240 ms (short RSI) or 540 ms (long RSI) was used. The flanker colors were presented first, and after another 50 ms the target color (S2) was displayed within S1, calling for the second response. Both flankers and target disappeared after 200 ms. There was no feedback for errors or omissions, and the next trial started after an intertrial interval of 800 ms.
To vary the expectedness of action effects, two variables were manipulated: First, the action effect E was either spatially compatible with the response (i.e., the lower key produces the bottom puzzle piece, the upper key produces the top puzzle piece) or spatially incompatible with the response (the lower key produces the top puzzle piece, the upper key produces the lower puzzle piece). Second, blocks consisted of either 75% compatible and 25% incompatible action effects (high-compatible blocks) or 25% compatible and 75% incompatible trials (high-incompatible blocks). Together, these two manipulations defined the variable expectedness. Unexpected action effects were therefore presented in the compatible trials of high-incompatible blocks and incompatible trials in high-compatible blocks, whereas the remaining trials presented expected action effects.
Participants started with three training blocks of 24 trials with the probe task only, to practice the response mapping in the probe task. In a fourth training block, the prime task was introduced and the two tasks were practiced together, with compatible action effects in all trials. After completion of the training blocks, recording of the electroencephalogram (EEG) started, and participants completed 26 blocks of 48 trials—13 blocks per block type (high-compatible vs. high-incompatible blocks), with each block containing 48 trials (36 expected action effects, 12 unexpected action effects). Here, participants were informed that puzzle pieces could also sometimes occur at the unexpected location. Block types switched after half of the session, and block type order (first half high-compatible vs. first half high-incompatible) was counterbalanced across participants.
Data recording
We recorded the EEG using a BIOSEMI Active-Two system (BioSemi, Amsterdam, The Netherlands) with 64 Ag–AgCl electrodes from channels Fp1, AF7, AF3, F1, F3, F5, F7, FT7, FC5, FC3, FC1, C1, C3, C5, T7, TP7, CP5, CP3, CP1, P1, P3, P5, P7, P9, PO7, PO3, O1, Iz, Oz, POz, Pz, CPz, Fpz, Fp2, AF8, AF4, AFz, Fz, F2, F4, F6, F8, FT8, FC6, FC4, FC2, FCz, Cz, C2, C4, C6, T8, TP8, CP6, CP4, CP2, P2, P4, P6, P8, P10, PO8, PO4, O2, as well as the left and right mastoid. The common mode sense and driven right leg electrodes were used as reference and ground electrodes. A vertical and horizontal electrooculogram (EOG) was recorded from electrodes above and below the right eye and on the outer canthi of both eyes. All electrodes were offline re-referenced to the averaged mastoids. EEG and EOG data were continuously recorded at a sampling rate of 1024 Hz and were resampled to 512 Hz offline.
Data analysis
Behavioral data
For the analysis of RTs, trials were excluded that deviated more than three standard deviations from the mean RT of each participant and condition (on average, 2.36% of trials). Response rates to the prime task and error rates to the probe task were arcsine-transformed for statistical testing to stabilize the variances (Winer, Brown, & Michels, 1991). RTs for the probe task were analyzed in a three-way repeated measures analysis of variance (ANOVA) on the variables prime expectedness (expected vs. unexpected), block type (high-compatible vs. high-incompatible), and probe correctness (correct vs. error). Error rates for the probe task were subjected to a two-way repeated measures ANOVA on the variables prime expectedness (expected vs. unexpected) and block type (high-compatible vs. high-incompatible). To simplify presentation of the results, the RSI was not considered in these analyses. However, to balance the contribution of each RSI to the means, all data were first averaged separately for each RSI and then averaged across the RSIs within each condition. Additional analyses that included the variable RSI did not reveal any interactions of this variable with either prime expectedness or block type.
Event-related potentials
We analyzed the EEG data using EEGLAB version 13.5 (Delorme & Makeig, 2004) and custom routines written in MatLab 8.3 (The Mathworks, Natick, MA).Footnote 2 First, the continuous EEG data were band-pass filtered, excluding activity below 0.1 Hz and above 40 Hz. Because we intended to examine neural activity with regard to error monitoring in the probe task, we extracted epochs that ranged from – 500 ms before to 1,000 ms after the response to the probe task. These epochs were baseline-corrected to – 150 to – 50 ms before response onset, to rule out any impacts of early onsets of the Ne/ERN. Subsequently, defective electrodes were interpolated using spherical spline interpolation if they met the joint probability criterion (threshold 5) as well as the kurtosis criterion (threshold 5) in EEGLAB’s channel rejection routine (pop_rejchan.m; M = 3.22 electrodes per participants). Epochs were removed if their amplitudes deviated more than 300 μV from zero or their joint probability deviated more than five standard deviations from the epoch mean (M = 77.93 trials per participant). An infomax-based independent components analysis was computed (Bell & Sejnowski, 1995) and components that represented ocular and muscular artifacts were removed after visual inspection. Grand averages were formed for all conditions and participants, and difference waves were computed from the grand averages of error trials minus those of correct trials. The conditions with the smallest trial numbers were errors. In these conditions, the following trial numbers resulted after artifact correction: high-compatible, expected—M = 62.7, SD = 22.7; high-compatible, unexpected—M = 23.3, SD = 9.2; high-incompatible, expected—M = 64.3, SD = 17.1; high-incompatible, unexpected—M = 19.0, SD = 4.8.
Both the Ne/ERN and the Pe were quantified in two ways that have been frequently reported in the literature on error monitoring: peak amplitudes in difference waves between correct and error trials (Falkenstein, Hoormann, Christ, & Hohnsbein, 2000; Holroyd & Coles, 2002) and peak-to-peak difference in error trials (Gehring et al., 1993; Nieuwenhuis et al., 2001). For the Ne/ERN, the peak amplitudes at electrode FCz were measured as the amplitude of the maximum difference between errors and correct responses (in the grand average) within 150 ms after response button press. Peak-to-peak amplitudes on error trials were quantified as the difference between the most positive peak within – 150 to 0 ms and the most negative peak within 150 ms after the response at electrode FCz (in the individual averages). Likewise, the Pe was quantified as the peak amplitude in the difference waves between 200 and 400 ms, and additionally as the peak-to-peak difference between the most negative peak within – 150 and 0 ms and the most positive peak within 200 to 400 ms, both at electrode Pz (Nieuwenhuis et al., 2001). All measures were analyzed using two-way repeated measures ANOVAs on the variables prime expectedness (expected vs. unexpected) and block type (high-compatible vs. high-incompatible). RSI was treated in the same way as for the behavioral analyses. Again, additional analyses including this variable did not reveal any significant interactions with prime expectedness or block type.
With regard to possible sensory attenuation after unexpected action effects (see Buzzell et al., 2017; Van der Borght et al., 2016), we analyzed the visual N1 in potentials that were time-locked to the onset of the flanker task stimulus, with a baseline period of – 200 to 0 ms before stimulus onset. Analogously to Buzzell et al., we identified a peak negativity at 120 ms over lateral-occipital electrodes, equally pronounced at PO7 and PO8, and the N1 was quantified as the mean amplitude during a 40-ms time window (100–140 ms) around this peak.
Results
Behavior
In the prime task, participants responded slightly more often with the bottom response (55.2%) than with the top response (44.8%), but this difference relied on only a few participantsFootnote 3 and was not significant, t(23) = 1.40, p = .174, d = 0.40. The associated mean RT was 283 ms.
The mean RTs for the probe task are presented in Fig. 2. Here we were interested in the impact of unexpected action effects in the prime task on the RTs of the probe task, since this was an indicator of the duration of the associated action effect monitoring (Wirth, Janczyk, & Kunde, 2018; Wirth, Steinhauser, et al., 2018). An ANOVA on the variables prime expectedness, block type, and probe correctnessFootnote 4 revealed that errors were faster (446 ms) than correct responses (474 ms), F(1, 23) = 16.1, p < .001, ηp2 = .41. Crucially, however, a main effect of prime expectedness, F(1, 23) = 4.89, p = .037, ηp2 = .18, showed that responses in the probe task were slower after unexpected action effects (463 ms) than after expected action effects (456 ms). Interestingly, we obtained a trend toward an interaction of prime expectedness and probe correctness, F(1, 23) = 3.35, p = .080, ηp2 = .13, indicating that the effect of prime expectedness—that is, the impact of effect monitoring—was more pronounced on error trials (unexpected: 452 ms vs. expected: 439 ms) than on correct trials (475 vs. 473 ms). This observation might have resulted because errors often reflect premature responses, which could be more susceptible to proactive interference from the action effect. In contrast, correct responses are based on more elaborate stimulus processing that may be better suited to overcome this interference. Nonetheless, this statistical trend (possibly due to a power limitation resulting from the sample size of N = 24) should be treated with caution. Taken together, these results demonstrate that increased monitoring for unexpected effects in the prime task slowed responses on the subsequent probe task, thus replicating our previous study (Wirth, Steinhauser, et al., 2018).

A Prime Expectedness × Block Type ANOVAFootnote 5 on the mean error rates of the probe task revealed a marginally significant main effect of block type, F(1, 23) = 3.96, p = .059, ηp2 = .15, which was qualified by a significant interaction between expectedness and block type, F(1, 23) = 15.4, p < .001, ηp2 = .40. Inspection of Fig. 2 reveals that this interaction mainly reflects that incompatible action effects (expected in high-incompatible blocks but unexpected in high-compatible blocks) led to higher error rates in the probe task than did compatible action effects.
ERP data
Consistent with the literature on error monitoring, grand averages of brain activity locked to the probe-task response showed a clear frontocentral Ne/ERN followed by a posterior Pe on error trials (Figs. 3 and 4). We first analyzed the Ne/ERN using two quantification procedures, each submitted to a Prime Expectedness × Block Type ANOVA. First, a peak-to-peak measure on error trials only revealed a significant main effect of prime expectedness, F(1, 23) = 11.5, p = .003, ηp2 = .33, indicating more negative Ne/ERN amplitudes after unexpected action effects (Fig. 3B). Second, a maximum peak analysis on the difference waves (correct minus error) yielded a significant main effect in the same direction (Fig. 3D), F(1, 23) = 5.03, p = .035, ηp2 = .18.Footnote 6

Grand averages (A) and difference waves (error minus correct, C) of frontocentral brain activity, locked to the response to the probe. Panels B and D show the Ne/ERN amplitudes for statistical testing, first based on a peak-to-peak analysis of the error trials (B), and second based on a minimum amplitude analysis of the difference waves (D). The thin lines in panel A represent correct trials. The scalp topography in panel C depicts the difference between correct and erroneous trials at 50 ms postresponse. Gray areas depict the time windows used for statistical testing.

Grand averages (A) and difference waves (error minus correct, C) of posterior brain activity, locked to the response to the probe. Panels B and D show the Pe amplitudes for statistical testing, first based on a peak-to-peak analysis of the error trials (B), and second based on a minimum amplitude analysis of the difference waves (D). The thin lines in panel A represent correct trials. The scalp topography in panel C depicts the difference between correct and erroneous trials at 300 ms postresponse. Gray areas depict the time windows used for statistical testing.
A similar pattern was obtained for the Pe (Fig. 4). The peak-to-peak analysis on error trials yielded a significant main effect of prime expectedness, F(1, 23) = 28.0, p < .001, ηp2 = .55, indicating larger Pe amplitudes after unexpected action effects than after expected action effects (Fig. 4B). Likewise, the peak amplitude analysis on difference waves confirmed this pattern (Fig. 4D), showing a significant main effect in the same direction, F(1, 23) = 9.07, p = .006, ηp2 = .28. Taken together, both quantification procedures demonstrated that both the Ne/ERN and the Pe were increased after unexpected action effects in the prime task. Please note on inspecting Figs. 3A and 4A that the Ne/ERN and Pe were quantified as peak-to-peak difference amplitudes, which may occur at slightly different latencies for each participant (mean latencies: Ne/ERN, M = 53 ms, SD = 24; PE, M = 323 ms, SD = 53). The visual impression of the averaged peak amplitudes in these figures may thus be misleading.
A last step of the analysis addressed the question of whether recent findings by Buzzell et al. (2017) and Van der Borght et al. (2016) on attenuated sensory processing after performance errors would generalize to trials following unexpected action effects. To this end, we investigated whether and how the visual N1 to the probe task, a neural correlate of early attentional processes, was affected by the expectancy of the preceding action effect.Footnote 7 Since Buzzell et al. suggested an involvement of the RSI in the N1 modulation, we subjected the flanker-stimulus-locked N1 data to an ANOVA on the variables prime expectedness, block type, and RSI (Fig. 5). In addition to the two main effects of expectedness, F(1, 23) = 6.83, p = .016, ηp2 = .23, and RSI, F(1, 23) = 14.9, p < .001, ηp2 = .39, an interaction of expectedness and RSI, F(1, 23) = 10.5, p = .004, ηp2 = .31, showed that the visual N1 was reduced after unexpected action effects only on short-RSI trials, F(1, 23) = 12.7, p = .002, ηp2 = .36, whereas no reduction could be observed on trials with a long RSI, F(1, 23) = 0.003, p = .96, ηp2 < .001.

Grand averages of lateral-occipital brain activity locked to the onset of the probe stimulus on trials with short (left) and long (right) response–stimulus intervals (RSIs). Gray areas depict the time window used for statistical testing.
Discussion
The goal of the present study was to investigate whether error monitoring and action effect monitoring rely on common mechanisms. Our central prediction was that activating the action effect monitoring system by means of unexpected action effects in a prime task should lead to enhanced monitoring of errors in a subsequent probe task. To this end, we adopted an experimental paradigm from previous studies in which increased monitoring of unexpected action effects has been demonstrated using behavioral and ERP measures (Wirth, Janczyk, & Kunde, 2018; Wirth, Steinhauser, et al., 2018). Our behavioral data replicated core aspects of these previous results: Presenting an unexpected action effect prolonged RTs for an immediately following probe task. This again demonstrated that action effect monitoring is a time-consuming process that interferes with subsequent processing, and that its duration varies with the expectedness of an action effect.
The crucial question in the present study was whether this enhanced action effect monitoring would generalize to error monitoring in the subsequent task. Analyses of the Ne/ERN and the Pe confirmed this prediction. Both the Ne/ERN and Pe to errors in the probe task were increased after unexpected action effects in the prime task. These modulations to error-related brain activity in the probe task occurred even though there was no experimental manipulation in this task itself. Instead, the Ne/ERN and Pe were exclusively influenced by changes to the preceding prime task, which strongly indicates a link between the monitoring systems in both tasks. Our results suggest that increased action effect monitoring for an unexpected effect in the prime task increases the sensitivity of subsequent error monitoring processes in the probe task.
A parsimonious explanation for this result is that both monitoring systems involve a mechanism that detects novel or unexpected outcomes. Preactivating this mechanism by means of unexpected action effects in the prime task implies that infrequent errors in the subsequent probe tasks are processed more strongly. This provides further support for the recent suggestion that the neural correlates of error monitoring at least partially reflect novelty detection rather than mere error- or reward-specific processing (Wessel et al., 2012). Wessel et al. (2012) found that similar neural activity underlies the Ne/ERN and a frontocentral novelty-N2 to frequency-matched oddball stimuli (see also Wessel & Aron, 2017). Our results are also consistent with the findings of Band et al. (2009), who found similar frontocentral negativities to task-irrelevant but infrequent action effects and negative feedback, that is, the FRN. This in turn allows our results to be linked to recent studies that suggest the FRN to represent processing of surprise and salience (Alexander & Brown, 2011; Hauser et al., 2014; Talmi, Atkinson, & El-Deredy, 2013), concepts closely related to the unexpectedness of the action effects in our paradigm.
Although these previous studies focused on frontocentral ERPs related to MFC activity, we observed that not only the Ne/ERN but also the Pe was enhanced following unexpected action effects. The Pe has frequently been assumed to be linked to the conscious awareness of an error (Nieuwenhuis et al., 2001; Overbeek et al., 2005). More specifically, one idea is that the Pe reflects an evidence accumulation process (M. Steinhauser & Yeung, 2010, 2012), receiving input from cognitive, autonomous, and sensory processing (Wessel et al., 2012; Wessel, Danielmeier, & Ullsperger, 2011). Provided that one possible input into this process is activity reflected by the Ne/ERN (Ullsperger et al., 2014; Ullsperger, Harsay, Wessel, & Ridderinkhof, 2010; Yeung et al., 2004), this could explain why effects on the Ne/ERN are carried over to the Pe in our data.
Another commonality that action effects and errors might share is negative affective valence. The Ne/ERN has previously been found to be susceptible to manipulations of affective valence (Larson, Perlstein, Stigge-Kaufman, Kelly, & Dotson, 2006; Wiswede, Münte, Goschke, & Rüsseler, 2009). Wiswede et al. presented emotional pictures shortly before a flanker task. The Ne/ERN on errors was increased after the presentation of unpleasant as compared to pleasant and neutral ones. The authors argued that this modulation of the Ne/ERN might originate in affect-driven changes to dopamine levels in the MFC, possibly in conjunction with an activation of the noradrenergic system due to increased arousal. Furthermore, spatial incompatibility entails response conflict (Simon, 1990) and recent evidence suggests that conflict and errors triggers negative affect (Aarts, De Houwer, & Pourtois, 2012, 2013; Dreisbach & Fischer, 2012; Maier, Scarpazza, Starita, Filogamo, & Làdavas, 2016; Schouppe et al., 2015; Wirth, Pfister, & Kunde, 2016). As a result, conflict elicited by unexpected action effects in our prime task could have generated negative affect, which in turn could have increased the sensitivity of detecting negatively valent errors in the probe task.
Irrespective of whether novelty or valence drives the present results, we believe that our results reflect changes in the sensitivity of monitoring due to the preactivation of the respective monitoring process. This is in line with previous studies that have indeed shown that the sensitivity of error monitoring can be influenced by contextual variables (e.g., the need to report errors, see Grützmann, Endrass, Klawohn, & Kathmann, 2014). Alternatively, one could assume that infrequent action effects prime errors. Priming would refer to a case in which the antecedent exposure to a prime stimulus enhances subsequent processing of a target that is identical to or associated with that prime (Bentin, McCarthy, & Wood, 1985; Meyer & Schvaneveldt, 1971). However, we think that priming is not an adequate concept to describe our data for two reasons: First, priming a stimulus can be considered as inducing an expectancy of that stimulus. However, this would imply that unexpected action effects increase the expectedness of an error, which should manifest in a reduced Ne/ERN. Second, and in accordance with this idea, ERP findings on perceptual priming of stimuli robustly report a suppression rather than an enhancement of brain activity in a primed target (e.g., Brown & Xiang, 1998; Gruber & Müller, 2002). Instead, in our data, infrequent action effects in the prime task increased the Ne/ERN in the probe task. This shows that our results are unlikely to be related to a priming mechanism. Rather than priming specific representations, our interpretation assumes that the monitoring process itself becomes more engaged in the prime task, which then generalizes to the probe task.
An alternative explanation, one that has always to be taken into account in error monitoring studies, is based on the link between the Ne/ERN and error frequency. It has repeatedly been shown that conditions associated with higher error rates are accompanied by lower Ne/ERN amplitudes (e.g., Fischer, Klein, & Ullsperger, 2017; Gehring et al., 1993; Jessup et al., 2010; Oliveira et al., 2007), although this relationship can be reversed when more frequent errors are associated with higher error significance (Maier, Di Pellegrino, & Steinhauser, 2012; Maier & Steinhauser, 2016). This link can be explained both by response conflict accounts (Danielmeier, Wessel, Steinhauser, & Ullsperger, 2009; Yeung et al., 2004) and by prediction error accounts (Holroyd & Coles, 2002) of the Ne/ERN. This explanation, however, cannot account for the pattern of Ne/ERN amplitudes in the present study, because error rates and Ne/ERN amplitudes in the probe task clearly follow different patterns. Whereas error rates vary mainly as a function of the compatibility of the action effect, the Ne/ERN exhibits—equal to our findings on the Pe—a main effect of expectancy only.
It is important to note that increased monitoring after unexpected action effects in the prime task did not lead to reduced error rates in the subsequent probe task. Hence, we cannot assume that this link between the two tasks is of a directly adaptive nature that seeks to avoid errors in the second task. Rather, as shown by Jentzsch and colleagues (Dudschig & Jentzsch, 2009; Jentzsch & Dudschig, 2009; see also Houtman & Notebaert, 2013), these very monitoring processes can be a source of errors in themselves—particularly with short RSIs (Buzzell et al., 2017; M. Steinhauser, Ernst, & Ibald, 2017). The link between monitoring processes in the two tasks that we observed in this study fits better with Wessel’s (2018) recent account on adaptivity in error processing. He suggests a nonspecific cascade of processing steps to be triggered by any unexpected action outcome. Only if a mendable source of the unexpected event is identifiable—which is not the case for the unexpected action effects of our prime task—adaptive adjustments to a subsequent task can be invoked (see also R. Steinhauser, Maier, & Steinhauser, 2017).
In contrast to the increased sensitivity of performance monitoring, visual attention was reduced after unexpected action effects, at least on trials with short RSI. This is in line with recent studies who found a corresponding reduction of early attentional processes after performance errors following short RSIs (Buzzell et al., 2017; Van der Borght et al., 2016). The attenuation of the visual N1 after unexpected action effects in the present study is important for three reasons. First, it provides additional evidence for the account that action effect monitoring and performance monitoring draw on the same processes. Apparently, performance errors and (unexpected) action effects elicit the same kind of adjustments to visual attention in the subsequent trial, an effect that also in our study was influenced by the RSI (see Buzzell et al., 2017). Second, the decreased N1 after unexpected action effects also rules out that the increased Ne/ERN and Pe result from increased general and unspecific arousal after unexpected action effects. In this case, the N1 (and other stimulus-locked components) would have shown a similar increase as the Ne/ERN and Pe. Finally, it strengthens previous results, indicating that ongoing action effect monitoring interferes with the perceptual processes of a subsequent task (Wirth, Janczyk, & Kunde, 2018, Exps. 2 and 3). With higher monitoring demand resulting in reduced visual attention, subsequent (visual) stimuli are identified and responded to later than following lower monitoring demand.
To sum up, our results showed that monitoring processes between subsequent tasks are linked to each other and increased action effect monitoring for one task can preactivate error monitoring processes in the subsequent task. This preactivation is likely to be rooted in a generic system of either novelty or affect processing that encompasses and connects tasks in close temporal succession. In this way, our findings contribute to understanding how tasks interact and influence each other in environments that require fast switching between the tasks or even require overlapping task execution. We could show that tasks in such scenarios not only influence each other through interference from competing task sets (Kiesel et al., 2010) or structural limitations to concurrent task execution (Meyer & Kieras, 1997; Pashler, 1994). Instead, monitoring processes to different tasks also interact with each other, since they are part of the same superordinate neural structures.
Author note
This work was supported by grants within Priority Program (SPP) 1772 from the German Research Foundation (Deutsche Forschungsgemeinschaft): R.S. and M.S. by Grant No. STE 1708/4-1, R.W. and W.K. by Grant No. KU 1964/11-1, and R.S. and R.W. together by an additional Research Excellence Grant, No. KI 1388/7-1. M.J.’s work is supported by Grant No. JA 2307/3-1, as well as by the Institutional Strategy of the University of Tübingen (ZUK63).
Notes
As compared to QWERTY keyboards, the positions of the letters Z and Y are switched in QWERTZ keyboards.
Data and analysis scripts are available from the corresponding author upon request.
Two participants preferred the bottom response, with about 90% of presses, considerably over the top response. Since they did not differ from the other participants with regard to their execution of the probe task (error rates, RTs), and can thus be considered to have been actively engaged in the experiment, we decided not to remove them from the subsequent analyses.
An additional ANOVA was conducted that also included the variable RSI (short vs. long). Since two participants did not have trials in all design cells, this analysis was conducted on a subsample of 22 participants. Besides main effects of prime expectedness, F(1, 21) = 5.93, p = .024, ηp2 = .22; probe correctness, F(1, 21) = 29.6, p < .001, ηp2 = .58; and RSI, F(1, 21) = 24.4, p < .001, ηp2 = .54; and a Probe Correctness × RSI interaction, F(1, 21) = 11.8, p = .002, ηp2 = .36, the ANOVA yielded no significant interactions that involved the variable RSI (all ps > .18). The Prime Expectedness × Probe Correctness interaction remained a statistical trend, F(1, 21) = 3.51, p = .075, ηp2 = .14.
Likewise, an additional ANOVA that included the variable RSI was also conducted on error rates. RSI contributed significantly only as a main effect, F(1, 23) = 17.8, p < .001, ηp2 = .44. All interactions with RSI were not significant (all ps > .31).
Literature suggests that a variant of the Ne/ERN occurs also on correct trials, termed the correct-related negativity (CRN; see, e.g., Vidal, Hasbroucq, Grapperon, & Bonnet, 2000). We can rule out that the observed effects also affect correct trials, on the basis of two arguments. First, if the same modulations would affect correct and error trials in a similar way, these effects would be cancelled out (or strongly reduced) in the difference waves, which is not the case. Second, analyzing the minimum amplitudes of correct trials 0–150 ms after the stimulus yields no significant differences with regard to prime expectedness or block type, all Fs < 0.806, all ps > .378, all ηp2s < .030.
To rule out that the observed expectedness effect on the N1 resulted from prolonged effects related to the prime task, we also analyzed the ERPs of the short-RSI condition at electrodes PO7/PO8, time-locked to the prime task response. We analyzed the mean amplitudes in a time window that preceded processing of the probe task stimulus—that is, from 0 to 200 ms after the prime task response. Here we observed no significant differences between the conditions, all Fs(1, 23) < 0.81, all ps > .37.
References
Aarts, K., De Houwer, J., & Pourtois, G. (2012). Evidence for the automatic evaluation of self-generated actions. Cognition, 124, 117–127. https://doi.org/10.1016/j.cognition.2012.05.009
Aarts, K., De Houwer, J., & Pourtois, G. (2013). Erroneous and correct actions have a different affective valence: Evidence from ERPs. Emotion, 13, 960–973. https://doi.org/10.1037/a0032808
Alexander, W. H., & Brown, J. W. (2011). Medial prefrontal cortex as an action-outcome predictor. Nature Neuroscience, 14, 1338–1344. https://doi.org/10.1038/nn.2921
Band, G. P. H., van Steenbergen, H., Ridderinkhof, K. R., Falkenstein, M., & Hommel, B. (2009). Action-effect negativity: Irrelevant action effects are monitored like relevant feedback. Biological Psychology, 82, 211–218.
Barcelo, F., Escera, C., Corral, M. J., & Periáñez, J. A. (2006). Task switching and novelty processing activate a common neural network for cognitive control. Journal of Cognitive Neuroscience, 18, 1734–1748. https://doi.org/10.1162/jocn.2006.18.10.1734
Bell, A. J., & Sejnowski, T. J. (1995). An information-maximization approach to blind separation and blind deconvolution. Neural Computation, 7, 1129–1159.
Bentin, S., McCarthy, G., & Wood, C. C. (1985). Event-related potentials, lexical decision and semantic priming. Electroencephalography and Clinical Neurophysiology, 60, 343–355.
Boldt, A., & Yeung, N. (2015). Shared neural markers of decision confidence and error detection. Journal of Neuroscience, 35, 3478–3484. https://doi.org/10.1523/JNEUROSCI.0797-14.2015
Brown, M. W., & Xiang, J. Z. (1998). Recognition memory: Neuronal substrates of the judgement of prior occurrence. Progress in Neurobiology, 55, 149–189.
Buzzell, G. A., Beatty, P. J., Paquette, N. A., Roberts, D. M., & McDonald, C. G. (2017). Error-induced blindness: Error detection leads to impaired sensory processing and lower accuracy at short response–stimulus intervals. Journal of Neuroscience, 37, 2895–2903.
Cardoso-Leite, P., Mamassian, P., Schütz-Bosbach, S., & Waszak, F. (2010). A new look at sensory attenuation: Action-effect anticipation affects sensitivity, not response bias. Psychological Science, 21, 1740–1745. https://doi.org/10.1177/0956797610389187
Cousineau, D. (2005). Confidence intervals in within-subject designs: A simpler solution to Loftus and Masson’s method. Tutorials in Quantitative Methods for Psychology, 1, 42–45.
Danielmeier, C., & Ullsperger, M. (2011). Post-error adjustments. Frontiers in Psychology, 2, 233. https://doi.org/10.3389/fpsyg.2011.00233
Danielmeier, C., Wessel, J. R., Steinhauser, M., & Ullsperger, M. (2009). Modulation of the error-related negativity by response conflict. Psychophysiology, 46, 1288–1298. https://doi.org/10.1111/j.1469-8986.2009.00860
Delorme, A., & Makeig, S. (2004). EEGLAB: An open source toolbox for analysis of single-trial EEG dynamics including independent component analysis. Journal of Neuroscience Methods, 134, 9–21. https://doi.org/10.1016/j.jneumeth.2003.10.009
Desantis, A., Roussel, C., & Waszak, F. (2014). The temporal dynamics of the perceptual consequences of action-effect prediction. Cognition, 132, 243–250.
Dreisbach, G., & Fischer, R. (2012). Conflicts as aversive signals. Brain and Cognition, 78, 94–98. https://doi.org/10.1016/j.bandc.2011.12.003
Dudschig, C., & Jentzsch, I. (2009). Speeding before and slowing after errors: Is it all just strategy? Brain Research, 1296, 56–62.
Elsner, B., Hommel, B., Mentschel, C., Drzezga, A., Prinz, W., Conrad, B., & Siebner, H. (2002). Linking actions and their perceivable consequences in the human brain. NeuroImage, 17, 364–372.
Falkenstein, M., Hohnsbein, J., Hoormann, J., & Blanke, L. (1991). Effects of crossmodal divided attention on late ERP components: II. Error processing in choice reaction tasks. Electroencephalography and Clinical Neurophysiology, 78, 447–455. https://doi.org/10.1016/0013-4694(91)90062-9
Falkenstein, M., Hoormann, J., Christ, S., & Hohnsbein, J. (2000). ERP components on reaction errors and their functional significance: A tutorial. Biological Psychology, 51, 87–107.
Fischer, A. G., Klein, T. A., & Ullsperger, M. (2017). Comparing the error-related negativity across groups : The impact of error- and trial-number differences. Psychophysiology, 54, 998–1009.
Folstein, J. R., & Van Petten, C. (2008). Influence of cognitive control and mismatch on the N2 component of the ERP: A review. Psychophysiology, 45, 152–170.
Gehring, W. J., Goss, B., Coles, M. G. H., Meyer, D. E., & Donchin, E. (1993). A neural system for error detection and compensation. Psychological Science, 4, 385–390. https://doi.org/10.1111/j.1467-9280.1993.tb00586.x
Gentsch, A., Ullsperger, P., & Ullsperger, M. (2009). Dissociable medial frontal negativities from a common monitoring system for self- and externally caused failure of goal achievement. NeuroImage, 47, 2023–2030.
Gruber, T., & Müller, M. M. (2002). Effects of picture repetition on induced gamma band responses, evoked potentials, and phase synchrony in the human EEG. Cognitive Brain Research, 13, 377–392.
Grützmann, R., Endrass, T., Klawohn, J., & Kathmann, N. (2014). Response accuracy rating modulates ERN and Pe amplitudes. Biological Psychology, 96, 1–7.
Hauser, T. U., Iannaccone, R., Stämpfli, P., Drechsler, R., Brandeis, D., Walitza, S., & Brem, S. (2014). The feedback-related negativity (FRN) revisited: New insights into the localization, meaning and network organization. NeuroImage, 84, 159–168.
Holroyd, C. B., & Coles, M. G. H. (2002). The neural basis of human error processing: Reinforcement learning, dopamine, and the error-related negativity. Psychological Review, 109, 679–708. https://doi.org/10.1037/0033-295X.109.4.679
Holroyd, C. B., Dien, J., & Coles, M. G. H. (1998). Error-related scalp potentials elicited by hand and foot movements: Evidence for an output-independent error-processing system in humans. Neuroscience Letters, 242, 65–68.
Hommel, B., Müsseler, J., Aschersleben, G., & Prinz, W. (2001). The Theory of Event Coding (TEC): A framework for perception and action planning. Behavioral and Brain Sciences, 24, 849–878, disc. 878–937. https://doi.org/10.1017/S0140525X01000103
Houtman, F., & Notebaert, W. (2013). Blinded by an error. Cognition, 128, 228–236.
Hughes, G., & Waszak, F. (2011). ERP correlates of action effect prediction and visual sensory attenuation in voluntary action. NeuroImage, 56, 1632–1640.
Iwanaga, M. I. O., & Nittono, H. (2010). Unexpected action effects elicit deviance-related brain potentials and cause behavioral delay. Psychophysiology, 47, 281–288.
James, W. (1890). The principles of psychology. New York, NY: Dover.
Janczyk, M., Durst, M., & Ulrich, R. (2017). Action selection by temporally distal goal states. Psychonomic Bulletin & Review, 24, 467–473.
Janczyk, M., & Lerche, V. (2018). A diffusion model analysis of the response–effect compatibility effect. Journal of Experimental Psychology: General. Advance online publication. https://doi.org/10.1037/xge0000430
Jentzsch, I., & Dudschig, C. (2009). Why do we slow down after an error? Mechanisms underlying the effects of posterror slowing. Quarterly Journal of Experimental Psychology, 62, 209–218.
Jessup, R. K., Busemeyer, J. R., & Brown, J. W. (2010). Error effects in anterior cingulate cortex reverse when error likelihood is high. Journal of Neuroscience, 30, 3467–3472.
Kiesel, A., Steinhauser, M., Wendt, M., Falkenstein, M., Jost, K., Philipp, A. M., & Koch, I. (2010). Control and interference in task switching—A review. Psychological Bulletin, 136, 849–874. https://doi.org/10.1037/a0019842
Larson, M. J., Perlstein, W. M., Stigge-Kaufman, D., Kelly, K. G., & Dotson, V. M. (2006). Affective context-induced modulation of the error-related negativity. NeuroReport, 17, 329–333.
Maier, M. E., Di Pellegrino, G., & Steinhauser, M. (2012). Enhanced error-related negativity on flanker errors: Error expectancy or error significance? Psychophysiology, 49, 899–908.
Maier, M. E., Scarpazza, C., Starita, F., Filogamo, R., & Làdavas, E. (2016). Error monitoring is related to processing internal affective states. Cognitive, Affective, & Behavioral Neuroscience, 16, 1050–1062. https://doi.org/10.3758/s13415-016-0452-1
Maier, M. E., & Steinhauser, M. (2016). Error significance but not error expectancy predicts error-related negativities for different error types. Behavioural Brain Research, 297, 259–267.
Melcher, T., Weidema, M., Eenshuistra, R. M., Hommel, B., & Gruber, O. (2008). The neural substrate of the ideomotor principle: An event-related fMRI analysis. NeuroImage, 39, 1274–1288.
Meyer, D. E., & Kieras, D. E. (1997). A computational theory of executive cognitive processes and multiple task performance: Part 1. Basic mechanisms. Psychological Review, 104, 3–65. https://doi.org/10.1037/0033-295X.104.1.3
Meyer, D. E., & Schvaneveldt, R. W. (1971). Facilitation in recognizing pairs of words: Evidence of a dependence between retrieval operations. Journal of Experimental Psychology, 90, 227–234. https://doi.org/10.1037/h0031564
Miltner, W. H. R., Braun, C. H., & Coles, M. G. H. (1997). Event-related brain potentials following incorrect feedback in a time-estimation task: Evidence for a “generic” neural system for error detection. Journal of Cognitive Neuroscience, 9, 788–798. https://doi.org/10.1162/jocn.1997.9.6.788
Morey, R. D. (2008). Confidence intervals from normalized data: A correction to Cousineau (2005). Tutorials in Quantitative Methods for Psychology, 4, 61–64.
Näätänen, R., & Gaillard, A. W. K. (1983). The orienting reflex and the N2 deflection of the event-related potential (ERP). In R. Näätänen & A. W. K. Gaillard (Eds.), Tutorials in ERP research endogenous components (pp. 119–141). Amsterdam, The Netherlands: North-Holland.
Naefgen, C., Dambacher, M., & Janczyk, M. (2017). Why free choices take longer than forced choices: Evidence from response threshold manipulations. Psychological Research. Advance online publication. https://doi.org/10.1007/s00426-017-0887-1
Nieuwenhuis, S., Ridderinkhof, K. R., Blom, J., Band, G. P. H., & Kok, A. (2001). Error-related brain potentials are differentially related to awareness of response errors: Evidence from an antisaccade task. Psychophysiology, 38, 752–760. https://doi.org/10.1111/1469-8986.3850752
Oliveira, F. T. P., McDonald, J. J., & Goodman, D. (2007). Performance monitoring in the anterior cingulate is not all error related: Expectancy deviation and the representation of action–outcome associations. Journal of Cognitive Neuroscience, 19, 1994–2004. https://doi.org/10.1162/jocn.2007.19.12.1994
Overbeek, T. J. M., Nieuwenhuis, S., & Ridderinkhof, K. R. (2005). Dissociable components of error processing: On the functional significance of the Pe vis-à-vis the ERN/Ne. Journal of Psychophysiology, 19, 319–329.
Pashler, H. (1994). Dual-task interference in simple tasks: Data and theory. Psychological Bulletin, 116, 220–244. https://doi.org/10.1037/0033-2909.116.2.220
Pfister, R., & Kunde, W. (2013). Dissecting the response in response–effect compatibility. Experimental Brain Research, 224, 647–655.
Polich, J. (2007). Updating P300: An integrative theory of P3a and P3b. Clinical Neurophysiology, 118, 2128–2148. https://doi.org/10.1016/j.clinph.2007.04.019
Polich, J., & Criado, J. R. (2006). Neuropsychology and neuropharmacology of P3a and P3b. International Journal of Psychophysiology, 60, 172–185.
Rabbitt, P., & Rodgers, B. (1977). What does a man do after he makes an error? an analysis of response programming. Quarterly Journal of Experimental Psychology, 29, 727–743. https://doi.org/10.1080/14640747708400645
Schouppe, N., Braem, S., De Houwer, J., Silvetti, M., Verguts, T., Ridderinkhof, K. R., & Notebaert, W. (2015). No pain, no gain: The affective valence of congruency conditions changes following a successful response. Cognitive, Affective, & Behavioral Neuroscience, 15, 251–261. https://doi.org/10.3758/s13415-014-0318-3
Shin, Y. K., Proctor, R. W., & Capaldi, E. J. (2010). A review of contemporary ideomotor theory. Psychological Bulletin, 136, 943–974. https://doi.org/10.1037/a0020541
Simon, J. R. (1990). The effects of an irrelevant directional cue on human information processing. In R. W. Proctor & T. G. Reeve (Eds.), Stimulus–response compatibility: An integrated perspective (pp. 31–86). Amsterdam, The Netherlands: North-Holland.
Steinhauser, M., Ernst, B., & Ibald, K. W. (2017). Isolating component processes of posterror slowing with the psychological refractory period paradigm. Journal of Experimental Psychology: Learning, Memory, and Cognition, 43, 653–659.
Steinhauser, M., & Kiesel, A. (2011). Performance monitoring and the causal attribution of errors. Cognitive, Affective, & Behavioral Neuroscience, 11, 309–320.
Steinhauser, M., & Yeung, N. (2010). Decision processes in human performance monitoring. Journal of Neuroscience, 30, 15643–15653. https://doi.org/10.1523/JNEUROSCI.1899-10.2010
Steinhauser, M., & Yeung, N. (2012). Error awareness as evidence accumulation: Effects of speed–accuracy trade-off on error signaling. Frontiers in Human Neuroscience, 6, 240:1–12. https://doi.org/10.3389/fnhum.2012.00053
Steinhauser, R., Maier, M. E., & Steinhauser, M. (2017). Neural signatures of adaptive post-error adjustments in visual search. NeuroImage, 150, 270–278.
Talmi, D., Atkinson, R., & El-Deredy, W. (2013). The feedback-related negativity signals salience prediction errors, not reward prediction errors. Journal of Neuroscience, 33, 8264–8269.
Ticini, L. F., Schütz-Bosbach, S., Weiss, C., Casile, A., & Waszak, F. (2012). When sounds become actions: Higher-order representation of newly learned action sounds in the human motor system. Journal of Cognitive Neuroscience, 24, 464–474.
Ullsperger, M., Fischer, A. G., Nigbur, R., & Endrass, T. (2014). Neural mechanisms and temporal dynamics of performance monitoring. Trends in Cognitive Sciences, 18, 259–267.
Ullsperger, M., Harsay, H. A., Wessel, J. R., & Ridderinkhof, K. R. (2010). Conscious perception of errors and its relation to the anterior insula. Brain Structure and Function, 214, 629–643.
Van der Borght, L., Schevernels, H., Burle, B., & Notebaert, W. (2016). Errors disrupt subsequent early attentional processes. PLoS ONE, 11, e0151843. https://doi.org/10.1371/journal.pone.0151843
van Veen, V., & Carter, C. S. (2002). The timing of action-monitoring processes in the anterior cingulate cortex. Journal of Cognitive Neuroscience, 14, 593–602. https://doi.org/10.1162/08989290260045837
Vidal, F., Hasbroucq, T., Grapperon, J., & Bonnet, M. (2000). Is the “error negativity” specific to errors? Biological Psychology, 51, 109–128.
Waszak, F., Cardoso-Leite, P., & Hughes, G. (2012). Action effect anticipation: Neurophysiological basis and functional consequences. Neuroscience & Biobehavioral Reviews, 36, 943–959.
Waszak, F., & Herwig, A. (2007). Effect anticipation modulates deviance processing in the brain. Brain Research, 1183, 74–82.
Welford, A. T. (1952). The “psychological refractory period” and the timing of high-speed performance—A review and a theory. British Journal of Psychology, 43, 2–19.
Wessel, J. R. (2018). An adaptive orienting theory of error processing. Psychophysiology, 55, 1–21.
Wessel, J. R., & Aron, A. R. (2017). On the globality of motor suppression: Unexpected events and their influence on behavior and cognition. Neuron, 93, 259–280.
Wessel, J. R., Danielmeier, C., Morton, J. B., & Ullsperger, M. (2012). Surprise and error: Common neuronal architecture for the processing of errors and novelty. Journal of Neuroscience, 32, 7528–7537. https://doi.org/10.1523/JNEUROSCI.6352-11.2012
Wessel, J. R., Danielmeier, C., & Ullsperger, M. (2011). Error awareness revisited: Accumulation of multimodal evidence from central and autonomic nervous systems. Journal of Cognitive Neuroscience, 23, 3021–3036. https://doi.org/10.1162/jocn.2011.21635
Wessel, J. R., Klein, T. A., Ott, D. V. M., & Ullsperger, M. (2014). Lesions to the prefrontal performance-monitoring network disrupt neural processing and adaptive behaviors after both errors and novelty. Cortex, 50, 45–54.
Winer, B. J., Brown, D. R., & Michels, K. M. (1991). Statistical principles in experimental design (3rd ed.). New York, NY: McGraw-Hill.
Wirth, R., Janczyk, M., & Kunde, W. (2018). Effect monitoring in dual-task performance. Journal of Experimental Psychology: Learning, Memory, and Cognition, 44, 553–571.
Wirth, R., Pfister, R., Brandes, J., & Kunde, W. (2016). Stroking me softly: Body-related effects in effect-based action control. Attention, Perception, & Psychophysics, 78, 1755–1770. https://doi.org/10.3758/s13414-016-1151-2
Wirth, R., Pfister, R., & Kunde, W. (2016). Asymmetric transfer effects between cognitive and affective task disturbances. Cognition and Emotion, 30, 399–416.
Wirth, R., Steinhauser, R., Janczyk, M., Steinhauser, M., & Kunde, W. (2018). Long-term and short-term action–effect links and their impact on effect monitoring. Journal of Experimental Psychology: Human Perception and Performance. Advance online publication. https://doi.org/10.1037/xhp0000524
Wiswede, D., Münte, T. F., Goschke, T., & Rüsseler, J. (2009). Modulation of the error-related negativity by induction of short-term negative affect. Neuropsychologia, 47, 83–90. https://doi.org/10.1016/j.neuropsychologia.2008.08.016
Yeung, N., Botvinick, M. M., & Cohen, J. D. (2004). The neural basis of error detection: Conflict monitoring and the error related negativity. Psychological Review, 11, 931–959. https://doi.org/10.1037/0033-295X.111.4.931
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Steinhauser, R., Wirth, R., Kunde, W. et al. Common mechanisms in error monitoring and action effect monitoring. Cogn Affect Behav Neurosci 18, 1159–1171 (2018). https://doi.org/10.3758/s13415-018-0628-y
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13415-018-0628-y