
Abstract
Meditation has been associated with relatively reduced activity in the default mode network, a brain network implicated in self-related thinking and mind wandering. However, previous imaging studies have typically compared meditation to rest, despite other studies having reported differences in brain activation patterns between meditators and controls at rest. Moreover, rest is associated with a range of brain activation patterns across individuals that has only recently begun to be better characterized. Therefore, in this study we compared meditation to another active cognitive task, both to replicate the findings that meditation is associated with relatively reduced default mode network activity and to extend these findings by testing whether default mode activity was reduced during meditation, beyond the typical reductions observed during effortful tasks. In addition, prior studies had used small groups, whereas in the present study we tested these hypotheses in a larger group. The results indicated that meditation is associated with reduced activations in the default mode network, relative to an active task, for meditators as compared to controls. Regions of the default mode network showing a Group × Task interaction included the posterior cingulate/precuneus and anterior cingulate cortex. These findings replicate and extend prior work indicating that the suppression of default mode processing may represent a central neural process in long-term meditation, and they suggest that meditation leads to relatively reduced default mode processing beyond that observed during another active cognitive task.
Similar content being viewed by others


Meditation involves maintaining attention on immediate experience and away from distractions such as self-referential thinking and mind wandering (Bishop et al., 2004). Consistent with this idea, meditation has been associated with relatively reduced activity in a network of brain regions implicated in self-referential processing, known as the default mode network (DMN), in experienced meditators relative to nonmeditators (Brewer et al., 2011b). Likewise, mind wandering has been associated with activity in the DMN (Mason et al., 2007), and reduced DMN activity during meditation has been associated with improved sustained attention outside of the scanner (Pagnoni, 2012). These findings suggest a role for reduced DMN processing during meditation.
Reduced DMN activity during meditation appears to be consistent across different meditation practices. A recent meta-analysis showed that DMN activity was consistently reduced during meditation, relative to control conditions, across neuroimaging studies of meditation that involved either focused attention or the repetition of phrases (Tomasino, Fregona, Skrap, & Fabbro, 2012). The same study by our research group revealed that DMN activity was reduced in meditators as compared to controls across three standard mindfulness meditations: focused concentration, loving kindness, and choiceless awareness (Brewer et al., 2011b). Determining that some neural mechanisms are common across meditation practices may inform the generalizability and potential clinical applications of these techniques.
The DMN has been found to be most highly active when individuals are left to think to themselves undisturbed or during tasks involving self-related processing, and less active during tasks requiring cognitive effort (Buckner, Andrews-Hanna, & Schacter, 2008; Raichle et al., 2001). This network is composed of a midline core, including the anterior medial prefrontal cortex and posterior cingulate cortex/precuneus; a dorsal medial prefrontal cortex subsystem including the temporal pole, lateral temporal cortex, and temporoparietal junction; and a medial temporal lobe subsystem including the ventral medial prefrontal cortex, posterior inferior parietal lobule, retrosplenial cortex, parahippocampal complex, and hippocampal formation (Andrews-Hanna, Reidler, Sepulcre, Poulin, & Buckner, 2010). Several of these brain regions, including the angular gyrus, middle temporal gyrus, and precuneus (Tomasino et al., 2012), have been shown across neuroimaging studies to have relatively reduced activity during meditation relative to control conditions, suggesting that increased cognitive effort and decreased self-related thinking are associated with meditation. In our prior study meditators showed lower activity during meditation than during rest in the posterior cingulate cortex and precuneus, relative to controls (Brewer et al., 2011b). Therefore, in the present study we aimed to replicate this finding with a larger sample, given that most neuroimaging studies of meditation—in particular, those involving experienced meditators—have used small groups (mean = 11.7, range = 4–31; Tomasino et al., 2012).
Previous studies have also reported that meditators, relative to controls, show differences in DMN activity not only during meditation, but also in functional connectivity at rest (Brewer et al., 2011b; Jang et al., 2011). These findings introduce a potential confound to studies of meditators that compare meditation to rest, because meditation may transform the resting state into a more meditative state. The choice of a control condition is a critical problem in cognitive neuroimaging studies and is fundamental for interpreting changes in brain activation patterns (Gusnard & Raichle, 2001; Marx et al., 2004). The resting brain state is expected to be highly variable across individuals, and therefore may be a poorer choice for comparison. To mitigate this confound, some studies have found it useful to compare meditation to active control tasks, such as mental arithmetic (e.g., Hölzel et al., 2007). Therefore, in this study we aimed to compare meditation to another active cognitive task, in order to test the hypothesis that meditation leads to reduced activity in the DMN beyond that found in another active cognitive task.
Method
Participants
All participants provided written informed consent in accordance with the Human Investigations Committee of the Yale School of Medicine. A total of 20 experienced meditators and 26 nonmeditators (controls) took part in the study. Of these participants, six meditators and three controls had participated in our previous study (Brewer et al., 2011b). All results reported here showed similar effects if the analyses were restricted to the new participants only. The meditators were recruited by advertisements and word of mouth and were all from the Insight meditation (Theravada) tradition. They reported a mean of 9,676 ± 1,586 practice hours over 14 ± 2 years, including daily practice and retreats. Controls reported no prior meditation experience. The groups were matched on sex, race, age, and years of education (Table 1).
fMRI protocol
Just before scanning, participants were instructed in three standard mindfulness meditation practices (as in previous studies: Brewer et al., 2011b; Gunaratana, 2002).
-
(a)
Concentration: “Please pay attention to the physical sensation of the breath wherever you feel it most strongly in the body. Follow the natural and spontaneous movement of the breath, not trying to change it in any way. Just pay attention to it. If you find that your attention has wandered to something else, gently but firmly bring it back to the physical sensation of the breath.”
-
(b)
Loving kindness: “Please think of a time when you genuinely wished someone well (pause). Using this feeling as a focus, silently wish all beings well, by repeating a few short phrases of your choosing over and over. For example: May all beings be happy, may all beings be healthy, may all beings be safe from harm.”
-
(c)
Choiceless awareness: “Please pay attention to whatever comes into your awareness, whether it is a thought, emotion, or body sensation. Just follow it until something else comes into your awareness, not trying to hold onto it or change it in any way. When something else comes into your awareness, just pay attention to it until the next thing comes along.”
Participants practiced each meditation condition outside of the scanner prior to fMRI and confirmed that they understood and could follow the instructions.
Each run began with a 30-s eyes-open rest period, during which participants were instructed to look at the fixation cross and not think of anything in particular. This was followed by an 8-s display of the instructions for the active cognitive task and by the 90-s active cognitive task itself. For the active task, participants were asked to make judgments about adjectives in response to a cue indicating that they should judge the word in terms of “self” (“Does the word describe you?”) or “case” (“Is the word in uppercase letters?”) and to indicate “yes” or “no” using a button box (Kelley et al., 2002). Adjectives were presented using E-Prime 1.2 (www.pstnet.com/eprime.cfm) for 2.5 s, with a 1- to 3-s interstimulus fixation interval for 30 trials per run, for a total of 180 trials. A total of 60 unique adjectives were drawn from the Anderson (1968) word list and were counterbalanced for valence. Participants practiced the active task to proficiency outside of the scanner prior to scanning. The active task was followed by a 30-s eyes-closed rest period. The eyes-closed condition was followed by a 30-s recorded meditation instruction (as above) and by a 180-s meditation period. At the end of the meditation period, subjects heard an audio prompt to open their eyes and rest until the sound of the scanner stopped, for an additional 20-s eyes-open rest period. Each meditation condition was performed twice, for a total of six runs. Meditation conditions were randomized, but the second instance of each meditation was blocked (i.e., AABBCC). After each run, participants were asked to rate how well they were able to follow the instructions and how much their mind wandered during meditation, on a scale from 0 to 10.
fMRI imaging parameters
Scanning was conducted using a Siemens 1.5-T Sonata MRI (Siemens AG, Erlangen, Germany) with an eight-channel receive-only head coil. High-resolution T1-weighted 3-D anatomical images were acquired using a magnetization-prepared rapid gradient echo (MPRAGE) sequence (TR = 2,530 ms, TE = 3.34 ms, field of view = 220 mm, matrix size = 192 × 192, slice thickness = 1.2 mm, flip angle = 8°, 160 slices). Low-resolution T1-weighted anatomical images were also acquired (TR = 500 ms, TE = 11 ms, field of view = 220 mm, slice thickness = 4 mm, gap = 1 mm, 25 AC–PC-aligned axial–oblique slices). Functional image acquisition began at the same slice location as in the T1 scan. Functional images were acquired using a T2*-weighted gradient-recalled single-shot echo-planar sequence (TR = 2,000 ms, TE = 35 ms, flip angle = 90°, bandwidth = 1446 Hz/pixel, matrix size = 64 × 64, field of view = 220 mm, voxel size = 3.5 mm, interleaved, 210 volumes; two volumes were acquired at the beginning of the run and discarded).
fMRI data preprocessing
Images were preprocessed using SPM8 (www.fil.ion.ucl.ac.uk/spm). The functional images were realigned for motion correction, and the resultant parameters were used as regressors of no interest in the fMRI model. In addition, Artifact Detection Tools (ART; www.nitrc.org/projects/artifact_detect) was used to identify global mean intensity and motion outliers in the fMRI time series (outlier thresholds: global signal > 3 standard deviations, motion > 1 mm). Any detected outliers were included as regressors of no interest in the model. A generative model of tissue classification, bias correction, and segmentation (Ashburner & Friston, 2005) was used to estimate the spatial normalization parameters to Montreal Neurological Institute (MNI) space. The estimates were then applied to all structural and functional images, and all images were smoothed using a 6-mm full-width-at-half-maximum Gaussian kernel.
Although motion outliers were modeled as regressors of no interest using ART, nonequivalent motion correction might result in bias when modeling group differences. Therefore, the mean outliers detected by ART across six runs were compared between groups using an independent t test. No significant difference in mean outliers was found between meditators and controls (meditators = 45, SEM = 6.3; controls = 38, SEM = 5.8), t(44) = 0.79, p = .43. Outliers were detected in all controls and in all but one meditator. Motion outliers in the first and last runs (Runs 1 and 6) were compared between the groups using a repeated measures analysis of variance. A significant effect of time was found (F = 4.34, p = .04), but no significant Group × Time interaction (F = 0.01, p = .91), such that the mean motion outliers increased from Runs 1 to 6 comparably in meditators (Run 1 = 5.1, Run 6 = 7.8) and controls (Run 1 = 5.9, Run 6 = 8.3).
fMRI data analysis
The blood oxygen level-dependent (BOLD) signal was modeled using separate regressors for the conditions: active task instructions, active task, meditation instructions, and meditation task. Rest periods were combined to form the implicit baseline. The meditation task included the three distinct meditation practices collapsed as blocks for the analysis. The active task included “self,” “case,” and fixation trials collapsed as blocks for the analysis. The conditions were modeled using a boxcar function convolved with a canonical hemodynamic response function, and the regressors were fit using SPM8’s implementation of the general linear model. To accommodate the long mediation conditions, the high-pass filter cutoff was 360 s. A first-level model was specified to estimate the parameter for each condition for each subject. A second-level model was specified to estimate the parameter for the main effects of task (meditation, active task) and group (meditation, control), and the interaction effect. A two-by-two interaction effect was tested using a repeated measures analysis of variance for groups (meditators, controls) by tasks (meditation, active task) and was exclusively masked with the group effect (meditation vs. control), in order to show the voxels in which the interaction was not driven by the main effect of group. All findings were significant at p ≤ .05 family-wise error (FWE) cluster-corrected, using a p ≤ .01 cluster-forming threshold and an extent threshold of 250 voxels, unless a more conservative threshold was indicated.
Statistics
The statistical analysis was conducted using SPSS 19 (http://www-01.ibm.com/software/analytic). For participant demographics, paired t tests were used to determine differences between the groups in age, and χ 2 tests were used to determine differences between the groups in sex. Repeated measures analyses of variance were used to determine differences between the groups in self-reported mind wandering. For the active task, independent t tests were used to compare reaction times between the groups, and χ 2 tests were used to compare error rates between the groups, with an error defined as an incorrect response to “case” or no response to “self.” All statistical tests were two-tailed and are reported as means ± standard deviations.
Results
Behavioral results
In line with the assumption that meditators and controls performed the active task similarly, no significant difference in reaction times was found between meditators (1.25 ± 0.38 s) and controls (1.26 ± 0.42 s), t = 1.46, p = .15. Meditators made significantly fewer errors in the “case” condition (1.7%) than did controls (3.5%), χ 2 = 13.2, p < .001, whereas no significant difference was found in errors in the “self” condition between groups (meditators = 1.7%, controls = 1.3%), χ 2 = 1.1, p = .31.
As expected, meditators reported less mind wandering during meditation than did controls, F(1, 44) = 7.57, p = .009. This finding was apparent for concentration (controls, 4.5 ± 2.1; meditators, 3.5 ± 1.4), loving kindness (controls, 3.8 ± 1.8; meditators, 2.8 ± 1.4), and choiceless awareness (controls, 4.4 ± 2.3; meditators, 2.7 ± 1.6) meditation. Both meditators and controls reported being able to follow the instructions to a high degree for concentration (controls, 8.6 ± 1.4; meditators, 8.5 ± 1.4), loving kindness (controls, 8.6 ± 1.4; meditators, 8.8 ± 1.2), and choiceless awareness (controls, 9.0 ± 1.4; meditators, 8.9 ± 0.9) meditation. No effect of time was found on mind wandering (meditators: Run 1, 3.0 ± 1.6; Run 6, 2.9 ± 2.0; controls: Run 1, 4.1 ± 2.0; Run 6, 4.3 ± 2.5), F(1, 44) = 0.003, p = .96, and likewise no Group × Time interaction was found for mind wandering, F(1, 44) = 0.19, p = .67. Similarly, no effect of time was found on the ability to follow instructions (meditators: Run 1, 8.7 ± 1.3; Run 6, 8.9 ± 1.4; controls: Run 1, 8.6 ± 1.2; Run 6, 8.7 ± 1.6), F(1, 44) = 1.14, p = .29, nor was a Group × Time interaction found for the ability to follow instructions, F(1, 44) = 0.57, p = .45.
fMRI results
For meditators and controls combined, meditation was associated with activity increases in the bilateral rectal gyrus and orbitofrontal cortex, relative to the implicit baseline (Fig. 1 top left, Table 2). The same brain regions showed an activity decrease during the active task relative to the implicit baseline in meditators and controls combined (Fig. 1 bottom right, Table 2).

Effects of task in the combined meditator and control groups. Meditation, as compared to the implicit baseline, is associated with activity increases bilaterally in the orbitofrontal cortex (top left). The same areas show an activity decrease during the active task, as compared to the implicit baseline (bottom right). Images are displayed in neurological convention, with critical thresholds at p < .001, uncorrected for multiple tests, to show the subthreshold extents of the effects.
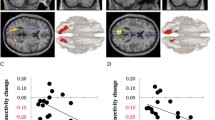
A between-group difference was found for meditation in comparison to the implicit baseline. Relative to controls, meditators showed reduced activity in the anterior cingulate cortex and the dorsal and ventral precuneus/posterior cingulate cortex during meditation, as compared to the implicit baseline (Fig. 2, supplementary Fig. S1).

A between-group contrast of meditation versus the implicit baseline revealed effects in the anterior cingulate cortex (ACC) and the dorsal (dPCu) and ventral precuneus (vPCu)/posterior cingulate cortex (M = meditators, C = controls). All three clusters are significant at p < .05 FWE-corrected, p ≤ .01 cluster-forming threshold, and extent threshold 250 voxels. Images are displayed in neurological convention, with critical thresholds at p < .01, uncorrected for multiple tests, to show the subthreshold extents of the effects.
A significant Group (meditators, controls) × Task (meditation, active task) interaction, exclusively masked by the effects of group, was identified in the middle temporal gyrus, fusiform and hippocampal gyri, anterior cingulate cortex, and precuneus (Fig. 3, Table 3). Plots of the parameter estimates for the anterior cingulate cortex and precuneus demonstrated that activity in these brain regions decreased during meditation and increased during the active control task in meditators, whereas controls did not show this dissociation (Fig. 3, insets).

A Group × Task interaction exclusively masked with the main effect of group revealed effects in the anterior cingulate cortex (ACC) and the dorsal precuneus (PCu) across groups and task conditions. Both clusters are significant at p < .05, FWE-corrected. Images are displayed in neurological convention, with critical thresholds of p < .01, uncorrected for multiple tests, to show the subthreshold extents of the effects. MM, meditators meditating; MA, meditators performing the active task; CM, controls meditating; CA, controls performing the active task.
Discussion
In this study, meditation was found to be associated with relatively lower activity in regions of the DMN in meditators than in controls, as compared to during another active cognitive task, indicated by a significant Group × Task interaction. Brain regions showing relatively reduced activity during meditation in meditators included the anterior cingulate cortex, fusiform gyrus, middle temporal gyrus, and precuneus. Meditators also showed relatively lower activity in DMN regions than did controls during meditation as compared to rest.
As we described above, the DMN is typically active during task-free resting states (Raichle et al., 2001), and this activity is thought to represent neural processing related to self-related thinking or mind wandering (Buckner et al., 2008). The DMN is further characterized by decreased activity during effortful, goal-directed tasks (Fox et al., 2005; Greicius, Krasnow, Reiss, & Menon, 2003). A recent meta-analysis reported that neuroimaging studies of meditation consistently report reduced DMN activity during meditation relative to control conditions in both meditators and nonmeditator controls (Tomasino et al., 2012). Although the meta-analysis did not find a difference in DMN activity associated with long-term experience, our prior study did show reduced activity in regions of the DMN during meditation relative to rest in experienced meditators compared with nonmeditators (Brewer et al., 2011b). This study replicated the results of that previous study in a larger sample (meditators, 20 vs. 12; controls, 26 vs. 12).
However, functional connectivity in regions of the DMN, a measure of the temporal correlation of the BOLD signal between these regions, has also been found to differ between meditators and controls, not only during meditation but also at rest (Brewer et al., 2011b; Pagnoni, 2012; Taylor et al., 2013). This suggests that meditation training may alter the behavioral state that individuals enter when given the standard resting-state instructions. Meditators and controls appear to differ in their resting-state DMN processing. Therefore, we compared meditation to another active cognitive task. Other studies have reported similar utility in comparing meditation with an active task (e.g., Hölzel et al., 2007; Tomasino et al., 2012). The present findings add to this work by providing evidence that meditation is associated with relatively reduced DMN activity during meditation as compared to a judgment-of-adjectives task in meditators versus controls. This finding suggests that meditation by experienced meditators leads to relatively reduced activity in the DMN, beyond that expected by general task-based deactivation.
Consistent with other prior findings (Kelley et al., 2002), our controls showed a pattern of reduced precuneus/posterior cingulate cortex activity during both the judgment-of-adjectives task and the meditation task (see the parameter estimate plots in the Figs. 2 and 3 insets). It is possible that for controls, reduced activity in this hub of the DMN during meditation and the active task reflects reduced self-related processing and mind wandering during these tasks in comparison with the implicit baseline, which was composed of resting periods. In support of this, task engagement has been shown to reduce activity in the precuneus/posterior cingulate cortex relative to rest (Fox et al., 2005). Other studies have reported a high incidence of mind wandering in healthy individuals (Killingsworth & Gilbert, 2010; Whitfield-Gabrieli et al., 2011) and a high incidence of precuneus/posterior cingulate cortex activity associated with mind wandering (Pagnoni, 2012). In contrast, meditators showed increased activity in the precuneus during the judgment-of-adjectives task (Fig. 2), possibly reflecting increased self-related processing relative to the implicit baseline. This interpretation would be consistent with our prior finding that meditators showed altered DMN functional connectivity at rest as compared to nonmeditators (Brewer et al., 2011b). Related to this, we used real-time fMRI neurofeedback, in which individuals were provided with dynamic visual feedback about their ongoing brain activity in real time, to demonstrate that the changes in activity in the posterior cingulate cortex corresponded to experienced meditators’ subjective reports of focused attention and mind wandering (Garrison et al., 2013a; Garrison et al., 2013b). The present findings further suggest that long-term meditation experience may lead to changes in DMN activity beyond typical task-engagement-related reductions, because meditators showed reduced DMN activity during meditation not only as compared to rest, but also as compared to another active cognitive task. For meditators, this is consistent with the hypothesis that meditation may reduce self-related thinking and mind wandering more than does another active task.
This study has several limitations. The use of a mixed design and the comparison of task blocks of different lengths may have reduced the design’s efficiency. Comparing blocks of different lengths can lead to poorer estimates of the shape of the hemodynamic response to a given stimulus block (Wager, Vazquez, Hernandez, & Noll, 2005). Block length was determined in consideration of both the task requirements and scan time limitations. To improve statistical power, the event-related active task (judgment of adjectives) was analyzed as a block. This might have combined events that increased (e.g., “self”) and decreased (e.g., “case”) DMN processing, thereby reducing power to detect DMN changes during this active task relative to meditation. Likewise, the meditation conditions (concentration, loving kindness, and choiceless awareness) were collapsed to improve power. This design could be optimized to directly compare the components of the active task and the different meditation practices in a future study. A related limitation was that the meditation and active tasks were not counterbalanced; the active task always preceded the meditation task. Although the fixed order was used to avoid specific effects of state-based meditation on brain activity patterns during the active task, this approach did not account for potential trait-based effects. Finally, interpretation of our results is limited to meditation in the research setting. Traditional or cultural meditation practices typically involve contextual components, such as intentions for practice, background conceptual beliefs, and the support of a community, among others. In the present study, meditation was performed in an fMRI scanner, and thus decontextualized. Despite these drawbacks, since the meditators were long-term practitioners with significant commitments to practice, we cannot rule out that larger components of the practice or memory of other contexts were active even during the decontextualized meditation tasks. Due to these empirical differences, further studies will be necessary to interpret our findings within the broader field of meditation research. Overall, despite the design limitations, this study showed reliable group differences in DMN activity across the different experimental conditions.
These findings provide evidence that reduced DMN processing may represent a central neural process in long-term meditation. This may have clinical implications. Previous work suggested that increased DMN activity may interfere with cognitive performance and that decreased DMN activity is associated with improved performance (for a review, see Anticevic et al., 2012). Likewise, increased DMN activity has been associated with depression (Sheline et al., 2009), anxiety (Zhao et al., 2007), and addiction (Garavan et al., 2000), among other disorders. Mind wandering and self-related processing contribute to ruminative thinking, which may be a feature of these disorders and has also been associated with decreased well-being (e.g., Killingsworth & Gilbert, 2010). In contrast, meditation, which appears to be associated with reduced activity in the DMN, has been shown to improve attention and working memory performance (Pagnoni, 2012) and promote positive health outcomes (Keng, Smoski, & Robins, 2011). Because mindfulness training has shown utility for addiction (Brewer et al., 2011a), as well as for pain, anxiety, and depression (Goyal et al., 2014), these studies together suggest that the neural mechanism by which meditation results in clinical benefits may be through reducing DMN activity.
References
Anderson, N. H. (1968). Likableness ratings of 555 personality-trait words. Journal of Personality and Social Psychology, 9, 272–279.
Andrews-Hanna, J. R., Reidler, J. S., Sepulcre, J., Poulin, R., & Buckner, R. L. (2010). Functional–anatomic fractionation of the brain’s default network. Neuron, 65, 550–562. doi:10.1016/j.neuron.2010.02.005
Anticevic, A., Cole, M. W., Murray, J. D., Corlett, P. R., Wang, X. J., & Krystal, J. H. (2012). The role of default network deactivation in cognition and disease. Trends in Cognitive Sciences, 16, 584–592. doi:10.1016/j.tics.2012.10.008
Ashburner, J., & Friston, K. J. (2005). Unified segmentation. NeuroImage, 26, 839–851. doi:10.1016/j.neuroimage.2005.02.018
Bishop, S. R., Lau, M., Shapiro, S., Carlson, L., Anderson, N. D., Carmody, J., . . . Devins, G. (2004). Mindfulness: A proposed operational definition. Clinical Psychology: Science and Practice, 11, 230–241. doi:10.1093/Clipsy/Bph077
Brewer, J. A., Mallik, S., Babuscio, T. A., Nich, C., Johnson, H. E., Deleone, C. M., . . . Rounsaville, B. J. (2011). Mindfulness training for smoking cessation: Results from a randomized controlled trial. Drug and Alcohol Dependence, 119, 72–80. doi:10.1016/j.drugalcdep.2011.05.027
Brewer, J. A., Worhunsky, P. D., Gray, J. R., Tang, Y. Y., Weber, J., & Kober, H. (2011b). Meditation experience is associated with differences in default mode network activity and connectivity. Proceedings of the National Academy of Sciences, 108, 20254–20259. doi:10.1073/pnas.1112029108
Buckner, R. L., Andrews-Hanna, J. R., & Schacter, D. L. (2008). The brain’s default network: Anatomy, function, and relevance to disease. In A. Kingstone & M. B. Miller (Eds.), The year in cognitive neuroscience 2008 (pp. 1–38). Malden, MA: Blackwell.
Fox, M. D., Snyder, A. Z., Vincent, J. L., Corbetta, M., Van Essen, D. C., & Raichle, M. E. (2005). The human brain is intrinsically organized into dynamic, anticorrelated functional networks. Proceedings of the National Academy of Sciences, 102, 9673–9678. doi:10.1073/pnas.0504136102
Garavan, H., Pankiewicz, J., Bloom, A., Cho, J. K., Sperry, L., Ross, T. J., . . . Stein, E. A. (2000). Cue-induced cocaine craving: Neuroanatomical specificity for drug users and drug stimuli. American Journal of Psychiatry, 157, 1789–1798.
Garrison, K. A., Santoyo, J. F., Davis, J. H., Thornhill, T. A. T., Kerr, C. E., & Brewer, J. A. (2013a). Effortless awareness: Using real time neurofeedback to investigate correlates of posterior cingulate cortex activity in meditators’ self-report. Frontiers in Human Neuroscience, 7, 440. doi:10.3389/fnhum.2013.00440
Garrison, K. A., Scheinost, D., Worhunsky, P. D., Elwafi, H. M., Thornhill, T. A., Thompson, E., . . . Brewer, J. A. (2013). Real-time fMRI links subjective experience with brain activity during focused attention. NeuroImage, 81, 110–118. doi:10.1016/j.neuroimage.2013.05.030
Goyal, M., Singh, S., Sibinga, E. M., Gould, N. F., Rowland-Seymour, A., Sharma, R., . . . Haythornthwaite, J. A. (2014). Meditation programs for psychological stress and well-being: A systematic review and meta-analysis. JAMA Internal Medicine, 174, 357–368. doi:10.1001/jamainternmed.2013.13018
Greicius, M. D., Krasnow, B., Reiss, A. L., & Menon, V. (2003). Functional connectivity in the resting brain: A network analysis of the default mode hypothesis. Proceedings of the National Academy of Sciences, 100, 253–258.
Gunaratana, H. (2002). Mindfulness in plain English. Somerville, MA: Wisdom Publications.
Gusnard, D. A., & Raichle, M. E. (2001). Searching for a baseline: Functionl imaging and the resting human brain. Nature Reviews Neuroscience, 2, 685–694.
Hölzel, B. K., Ott, U., Hempel, H., Hackl, A., Wolf, K., Stark, R., & Vaitl, D. (2007). Differential engagement of anterior cingulate and adjacent medial frontal cortex in adept meditators and non-meditators. Neuroscience Letters, 421, 16–21. doi:10.1016/j.neulet.2007.04.074
Jang, J. H., Jung, W. H., Kang, D.-H., Byun, M. S., Kwon, S. J., Choi, C.-H., & Kwon, J. S. (2011). Increased default mode network connectivity associated with meditation. Neuroscience Letters, 487, 358–362. doi:10.1016/j.neulet.2010.10.056
Kelley, W. M., Macrae, C. N., Wyland, C. L., Caglar, S., Inati, S., & Heatherton, T. F. (2002). Finding the self? An event-related fMRI study. Journal of Cognitive Neuroscience, 14, 785–794.
Keng, S. L., Smoski, M. J., & Robins, C. J. (2011). Effects of mindfulness on psychological health: A review of empirical studies. Clinical Psychology Review, 31, 1041–1056. doi:10.1016/j.cpr.2011.04.006
Killingsworth, M. A., & Gilbert, D. T. (2010). A Wandering Mind Is an Unhappy Mind. Science, 330, 932. doi:10.1126/science.1192439
Marx, E., Deutschlander, A., Stephan, T., Dieterich, M., Wiesmann, M., & Brandt, T. (2004). Eyes open and eyes closed as rest conditions: Impact on brain activation patterns. NeuroImage, 21, 1818–1824. doi:10.1016/j.neuroimage.2003.12.026
Mason, M. F., Norton, M. I., Van Horn, J. D., Wegner, D. M., Grafton, S. T., & Macrae, C. N. (2007). Wandering minds: The default network and stimulus-independent thought. Science, 315, 393–395. doi:10.1126/science.1131295
Pagnoni, G. (2012). Dynamical properties of BOLD activity from the ventral posteromedial cortex associated with meditation and attentional skills. Journal of Neuroscience, 32, 5242–5249. doi:10.1523/JNEUROSCI.4135-11.2012
Raichle, M. E., MacLeod, A. M., Snyder, A. Z., Powers, W. J., Gusnard, D. A., & Shulman, G. L. (2001). A default mode of brain function. Proceedings of the National Academy of Sciences, 98, 676–682. doi:10.1073/pnas.98.2.676
Sheline, Y. I., Barch, D. M., Price, J. L., Rundle, M. M., Vaishnavi, S. N., Snyder, A. Z., . . . Raichle, M. E. (2009). The default mode network and self-referential processes in depression. Proceedings of the National Academy of Sciences, 106, 1942–1947. doi:10.1073/pnas.0812686106
Taylor, V. A., Daneault, V., Grant, J., Scavone, G., Breton, E., Roffe-Vidal, S., . . . Beauregard, M. (2013). Impact of meditation training on the default mode network during a restful state. Social Cognitive and Affective Neuroscience, 8, 4–14. doi:10.1093/scan/nsr087
Tomasino, B., Fregona, S., Skrap, M., & Fabbro, F. (2012). Meditation-related activations are modulated by the practices needed to obtain it and by the expertise: An ALE meta-analysis study. Frontiers in Human Neuroscience, 6, 346. doi:10.3389/fnhum.2012.00346
Wager, T. D., Vazquez, A., Hernandez, L., & Noll, D. C. (2005). Accounting for nonlinear BOLD effects in fMRI: Parameter estimates and a model for prediction in rapid event-related studies. NeuroImage, 25, 206–218. doi:10.1016/j.neuroimage.2004.11.008
Whitfield-Gabrieli, S., Moran, J. M., Nieto-Castañón, A., Triantafyllou, C., Saxe, R., & Gabrieli, J. D. E. (2011). Associations and dissociations between default and self-reference networks in the human brain. NeuroImage, 55, 225–232. doi:10.1016/j.neuroimage.2010.11.048
Zhao, X. H., Wang, P. J., Li, C. B., Hu, Z. H., Xi, Q., Wu, W. Y., & Tang, X. W. (2007). Altered default mode network activity in patient with anxiety disorders: An fMRI study. European Journal of Radiology, 63, 373–378. doi:10.1016/j.ejrad.2007.02.006
Author note
This work was supported by awards from the National Institutes of Health, National Institute on Drug Abuse (Grant No. K12-DA00167 to J.A.B... and K.A.G.); from the US Veterans Affairs New England Mental Illness Research, Education, and Clinical Center; and from the American Heart Association (Grant No. 14CRP18200010 to K.A.G.), and by private donations from Jeffrey C. Walker, Austin Hearst, and the 1440 Foundation. We thank our participants for their time and effort, Joseph Goldstein and Ginny Morgan for input on the meditation instructions, Hedy Kober for input on the study design, Thomas Thornhill IV for study coordination, and Hedy Sarofin and the staff of the Yale Magnetic Resonance Research Center for help with scanning.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Supplemental Fig. S1
Effects of group for meditators during meditation (top row) and during the active task (bottom row). Images are displayed in neurological convention, with critical thresholds at p < .01, uncorrected for multiple tests, to show the subthreshold extents of the effects. (JPEG 177 kb)
Rights and permissions
About this article

{kind=link}
Cite this article
Garrison, K.A., Zeffiro, T.A., Scheinost, D. et al. Meditation leads to reduced default mode network activity beyond an active task. Cogn Affect Behav Neurosci 15, 712–720 (2015). https://doi.org/10.3758/s13415-015-0358-3
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13415-015-0358-3