
Abstract
Botvinick, Cognitive, Affective, & Behavioral Neuroscience 7:356–366 (2007) recently suggested that competing theories of the monitoring function of the anterior cingulate cortex (ACC) for cognitive control might converge on the detection of aversive signals in general, implying that response conflicts, a known trigger of ACC activation, are aversive, too. Recent evidence showing conflict priming (i.e., faster responses to negative targets after conflict primes) directly supports this notion but remains inconclusive with regard to possible confounds with processing fluency. To this end, two experiments were conducted to offer more compelling evidence for the negative valence of conflicts. Participants were primed by (conflict and nonconflict) Stroop stimuli and subsequently had to judge the valence of neutral German words (Experiment 1a) or Chinese pictographs (Experiment 1b). Results showed that conflict, as compared with nonconflict, primes led to more negative judgments of subsequently presented neutral target stimuli. The findings will be discussed in the light of existing theories of action control highlighting the role of aversive signals for sequential processing adjustments.
Similar content being viewed by others

Cognitive control describes the human ability to flexibly adapt goals and actions in accordance with internal and external needs (Miller & Cohen, 2001). One of the main mechanisms underlying such cognitive flexibility is a monitoring function that supervises the ongoing processing stream for significant information—such as, for example, error feedback or coactivation of conflicting response tendencies. Once an error or conflict signal is detected, cognitive control is increased in order to solve the conflict or increase accuracy, respectively (e.g., Botvinick, Braver, Barch, Carter, & Cohen, 2001; Holroyd & Cole, 2002; Yeung, Botvinick, & Cohen, 2004). From brain imaging studies, there exists broad empirical evidence that the anterior cingulate cortex (ACC) plays a major role in this monitoring process. For example, Botvinick and colleagues (2001), in their influential conflict monitoring theory, suggest that the ACC is activated by response conflicts and then signals the need for additional control to the dorsolateral prefrontal cortex (Kerns et al., 2004; MacDonald, Cohen, Stenger, & Carter, 2000). Behaviorally, this sequential control adaptation is reflected in a reduced response conflict on trial N after a response conflict on trial N − 1 (Botvinick, Nystrom, Fissell, Carter, & Cohen, 1999; Gratton, Coles, & Donchin, 1992; Notebaert, Soetens, & Melis, 2001; Stürmer, Leuthold, Soetens, Schröter, & Sommer, 2002; Wühr & Ansorge, 2005; for a review, see Egner, 2007). However, this ACC model of conflict adaptation was recently challenged by several studies showing that the ACC is activated not only by response conflicts, but also by social rejection/exclusion (Eisenberger, Lieberman, & Williams, 2003), monetary losses (Nieuwenhuis, Yeung, Holroyd, Schurger, & Cohen, 2004), and experienced and witnessed pain (Rainville, 2002; Singer et al., 2004; for a review, see Shackman et al., 2011). Therefore, Botvinick (2007) presented an integrative account of ACC function, suggesting that the ACC might monitor and detect just any aversive signal in the ongoing processing stream. The significance of this assumption should not be underestimated, since it would make the ACC monitoring theory applicable to a much wider range of tasks and situations. However, since most of the evidence in favor of the monitoring theory of ACC function stems from response conflict paradigms, it is essential to show that response conflicts themselves actually serve as an aversive signal.
The notion of conflict as an aversive signal so far has mostly been supported by indirect evidence. Using an Eriksen flanker paradigm, van Steenbergen, Band, and Hommel (2009) provided the first indirect evidence for the aversive nature of conflicts. They showed that reward stimuli randomly (and not conditional on the actual performance) following conflict trials eliminated conflict adaptation effects. This was taken as evidence that the positive affect induced by the reward signal counteracted the negative valence of the conflict, thereby eliminating the signal for control adjustments (but see Braem, Verguts, Roggeman, & Notebaert, 2012, who found enhanced conflict adaptation following action-contingent reward). Furthermore, Dreisbach and Fischer (2011) provided support that the negative valence of conflicts, and not the response conflict itself, triggers sequential processing adjustments. In a magnitude comparison task with no response conflict involved, participants had to decide whether a presented number word was smaller or larger than five. The number words were written in either an easy-to-read (fluent) or a hard-to-read (nonfluent) font. It is well-documented that stimuli that are processed fluently induce positive affect, whereas stimuli that are processed nonfluently induce negative affect (e.g., Reber et al., 2004; Winkielman & Cacioppo, 2001). Importantly, the results presented by Dreisbach and Fischer (2011) show that nonfluent and, thus, aversive number words induced sequential processing adjustments resembling conflict adaptation effects: The fluency effect (faster responses for fluent than for nonfluent trials) was smaller following nonfluent trials than following fluent trials. This was taken as evidence that even in the absence of conflict, cognitive effort increased after nonfluent stimuli. Combining literature that highlights the aversiveness of nonfluent stimuli with the observation that they induce conflict-like sequential adaptation effects, it can thus be reasoned that it is the aversiveness inherited in both types of stimuli that triggers the control adjustments. Further evidence for the aversiveness of conflict comes from Schouppe, De Houwer, Ridderinkhof, and Notebaert (2012), who recently showed that Stroop conflict stimuli promote avoidance behavior. Probably the most direct approach to studying the affective valence of conflicts was taken by Dreisbach and Fischer (2012a), who administered a conflict priming task, inspired by the affective priming paradigm first introduced by Fazio, Sanbonmatsu, Powell, and Kardes (1986). In a typical affective priming paradigm, participants encounter a prime stimulus of a certain valence (e.g., a positive or negative picture), followed by a target stimulus of a certain valence (e.g., a positive or negative word), with the instruction to evaluate the valence of the target stimulus as quickly and accurately as possible. The typical finding is that positive primes facilitate responses to positive targets, whereas negative primes facilitate responses to negative targets. Now Dreisbach and Fischer (2012a) used congruent and incongruent Stroop color words as (presumably) affective primes and positive and negative words/pictures as targets. As was predicted, they found shorter reaction times (RTs) for negative targets following incongruent Stroop primes and longer RTs following congruent Stroop primes. This was taken as the first direct evidence for the negative valence of conflicts. However, and alternatively, the results might also be interpreted in terms of a match of processing fluency between the prime and target, as will be further outlined below. Therefore, the aim of the study presented here was to provide more unequivocal evidence for the aversive nature of conflicts.
Affective priming effects are commonly referred to as effects of valence compatibility (Fazio, 2001; Fazio et al., 1986): Responses are slow whenever the valences of the prime and target are incompatible and fast whenever both valences are compatible. However, it is conceivable that conflict primes and negative targets in the study by Dreisbach and Fischer (2012a, b) matched in more than just the valence dimension. The numerous variants of priming paradigms suggest that the possibilities for an overlap in dimensions and, thus, compatibility effects between the prime and target stimuli are abundant (see Kornblum, Hasbroucq, & Osman, 1990). For instance, the prime and target can overlap semantically, such as dog priming cat instead of sun, or orthographically, such as house priming mouse instead of rose (for a review, see Gulan & Valerjev, 2010). Importantly, findings by Chang and Mitchell (2009) suggest that results in compatibility-dependent tasks with more than one dimensional overlap between the prime and target can be confounded with unnoticed prime–target relations. More specifically, the authors report that findings in the implicit association task (Greenwald, McGhee, & Schwartz, 1998), an associative task closely related to affective priming paradigms, can be confounded with artifacts due to underlying compatibility in terms of processing fluency. Perhaps, then, faster responses in nonconflict-prime/positive-target and conflict-prime/negative-target combinations (Dreisbach & Fischer, 2012a) were not the result of valence associations but were caused by matching processing characteristics: In particular, positive targets and nonconflict primes might be processed more easily, and negative targets and conflict primes might be processed less easily (see Scott, O’Donnell, Leuthold, & Sereno, 2009; Unkelbach, Fiedler, Bayer, Stegmüller, & Danner, 2008). One theoretical account, the so-called density hypothesis, proposes that the speed advantage in the processing of positive information, as compared with negative information, is caused by the higher densitiy of positive information in memory (i.e., positive concepts such as “happy” and “content” are more similar than negative concepts such as “angry” and “sad,” for example; Unkelbach et al., 2008). Because of this alleged density, processing of a positive stimulus (in comparison with processing of a negative stimulus) immediately triggers an entire network of equally valenced information. This fast and widespread activation manifests in higher processing fluency for positive information, whereas the processing of negative information triggers a rather uncompressed network, manifesting in lower processing fluency for negative information. Applied to the conflict paradigm used previously, this means that priming effects between nonfluently processed negative targets and nonfluently processed incongruent Stroop stimuli (as compared with congruent Stroop stimuli) might rest on similar processing characteristics instead of valence congruency. That is, even though affect and processing fluency may represent two sides of the same coin (Reber et al., 2004; Winkielman & Cacioppo, 2001), it cannot be ruled out that the priming effects observed by Dreisbach and Fischer (2012a) were mediated mainly by an overlap in processing fluency between the conflict prime and target. To this end, in the experiments presented here, we again administered a conflict priming task, but this time used only neutral targets to deconfound possible fluency characteristics from target valence. Participants’ task this time was to spontaneously judge the affective valence of the neutral target stimuli. The rationale is that conflict primes should modulate the affective judgment according to the prime’s valence (e.g., Murphy, Monahan & Zajonc, 1995; Payne, Cheng, Govorun, & Stewart, 2005). Whereas RTs for categorizing affective target stimuli according to their valence served as the dependent measure in the conflict priming paradigm used by Dreisbach and Fischer (2012a), we now measured valence judgments of affectively neutral target stimuli (i.e., German words in Experiment 1a and neutral Chinese characters in Experiment 1b) following congruent and incongruent Stroop primes. That is, neutral target stimuli ensured equivalent processing fluency of targets and, thus, ruled out the possibility of fluency compatibility confounds. If conflicts are truly aversive in nature, their negative valence should spread to the otherwise neutral targets when presented shortly after them. We thus expected to find more negative judgments after conflict primes than after nonconflict primes.
Experiments 1a and 1b
Method
Participants
In Experiment 1a, 21 right-handed students (13 female, eight male; mean age = 25.6 years, SD = 5.2) and, in Experiment 1b, a separate sample of 24 right-handed students (19 female, five male; mean age = 22.2 years, SD = 1.8) from the University of Regensburg participated in exchange for a chocolate bar or 2 Euros. All 45 participants signed informed consent and were debriefed after the session. The data of 1 participant who showed a strong trend toward positive judgment (more than 75 % of target words were evaluated as positive; Experiment 1a), 1 participant who was color blind (Experiment 1b), and 1 participant who misunderstood instructions (Experiment 1b) were excluded from analysis, leaving final samples of 20 (Experiment 1a) and 22 (Experiment 1b).
Apparatus and stimuli
Primes were the German color words for BLUE, GREEN, YELLOW, RED, and PURPLE printed in blue, green, yellow, red, and purple. The print color could either match (nonconflict stimuli) or mismatch (conflict stimuli) the color denoted by the word. Primes and target words were written in Courier New bold, 26 point, each letter subtending a visual angle of approximately 0.7° × 0.7° at a viewing distance of 55 cm. In Experiment 1a, targets were 112 German words (94 nouns, 12 verbs, and 6 adjectives selected on the basis of neutral affective ratings from the Berlin affective word-list reloaded BAWL–R (mean valence ratings: −.001, running on a scale from −3 [very negative] through 0 [neutral ] to 3 [very positive], SD = .000; range of ratings: .109; Võ et al., 2009). In Experiment 1b, targets were 144 Chinese pictographs that subtended a visual angle of approximately 8° × 8° and were chosen randomly from an online English–Chinese dictionaryFootnote 1 (cf. Murphy et al., 1995; Murphy & Zajonc, 1993; Zajonc, 1968). The pictographs represented nouns, such as wall, verbs, such as to go, and adjectives, such as new. Primes and targets appeared at the center of the screen on a light gray background.
Procedure
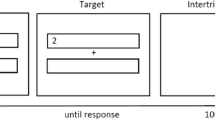
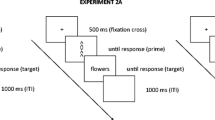
Participants had to judge the valence of targets (i.e., German words [Experiment 1a] or Chinese characters [Experiment 1b]) that were preceded by Stroop primes. They had to press a right response key to judge the target as positive and a left response key to judge the target as negative (“-“ and “y” keys on a QWERTZ keyboard, respectively). This assignment was held constant, since people have a natural tendency to associate positive concepts with their dominant body side—that is, the right hand (Casasanto, 2009). Participants were asked to choose each option at approximately the same rate to minimize potential biases in favor of one response key. To ensure processing of the primes, catch trials were interspersed: Whenever the prime word was the German word for PURPLE or the primes BLUE, GREEN, YELLOW, or RED were printed in purple, participants had to press the space bar instead of evaluating the following target. Since both color and word dimension could denote a potential catch trial, proper encoding of Stroop primes and, thus, the experience of conflict should be warranted.
Each trial started with the presentation of the Stroop prime for 400 ms, since electrophysiological studies have shown a peak in conflict-associated negativity at around 400 ms after Stroop stimulus onset (Hanslmayr et al., 2008; Liotti, Woldorff, Perez, & Mayberg, 2000). After the prime, the target appeared and remained on the screen until a response was given. Intertrial intervals were of 1,000-ms length. On erroneous catch-trials, the German word for ERROR appeared on the screen.
After a short test to exclude color blindness, the conflict priming task started with two short practice blocks to familiarize participants with the task.Footnote 2 Experiment 1a consisted of two experimental blocks of 112 trials, separated by a self-paced break. In a given block, each target word was presented once. More precisely, 48 words were preceded by a congruent prime, 48 by an incongruent prime, and 16 by a catch prime in a given block. Prime congruency was thus manipulated trial-wise and was random. Moreover, target words that were preceded by a conflict prime in one block were preceded by a nonconflict prime in the second block (order counterbalanced across participants). Experiment 1b, using 144 different Chinese characters, consisted of only one experimental block of 144 trials length, composed of 120 target trials intermixed with 24 catch trials. Sixty targets were thus preceded by a congruent prime, 60 targets by an incongruent prime, and 24 by a catch prime. Again, the combination of prime congruence and target was random and varied between participants. In both experiments, there was a minimum distance of 2 and a maximum distance of 8 target trials between catch trials. Experiment 1a lasted about 15 min, Experiment 1b about 8 min.
Results
Mean error rate on catch trials was low in both experiments (1a, M = 4.08 %, SD = 3.68; 1b, 4.91 %, SD = 7.41), suggesting that participants encoded both word and color dimension properly. For each experiment, proportions of negative/positive judgments as a function of prime (conflict/nonconflict) in the experimental blocks were computed. To test for a general bias in judgment, we conducted two one-sample t-tests (two-sided) against the null hypothesis of 50 %. In both experiments, judgment was unbiased [1a, frequency of positive judgments, M = 52.37 %, SD = 8.56, t(19) = 1.239, p = .231, d = 0.281; 1b, M = 48.58 %, SD = 8.13, t(21) < 1, p = .423, d = 0.183].Footnote 3
To test for affective priming effects by conflict primes, we compared mean frequencies of negatively judged targets depending on prime condition. In both experiments, paired t-tests (one-sided) revealed a significant difference between prime conditions [1a, t(19) = 2.125, p = .024, d = 0.475; 1b, t(21) = 2.256, p = .018, d = 0.481]. Neutral target stimuli were judged more frequently as negative after conflict than after nonconflict primes (1a, M = 48.54 %, SD = 9.30, vs. M = 46.32 %, SD = 8.95; 1b, M = 53.52 %, SD = 9.35, vs. M = 49.31 %, SD = 9.11; see Fig. 1).

Frequencies (in percentages) of negative judgments of neutral targets as a function of prime condition (nonconflict, conflict) in Experiments 1a and 1b. Bars represent standard errors of the means
Discussion
The results presented here clearly support the notion of conflicts as aversive signals as implied by the integrative account of ACC function recently suggested by Botvinick (2007). The negative valence of conflicts, just like the negative affect induced by frowning faces (Murphy & Zajonc, 1993) or negative pictures (Payne, Shimizu, & Jacoby, 2005), spreads to neutral target stimuli that are then evaluated accordingly: Neutral German words (Experiment 1a) and Chinese pictographs (Experiment 1b) were more often judged as negative after conflict than after nonconflict primes. Whereas findings of conflict priming in paradigms relying on prime–target compatibility (Dreisbach & Fischer, 2012a) were ambiguous with regard to their origin (i.e., match vs. mismatch of either valence or processing fluency between the prime and target), the two experiments presented here provide unequivocal evidence for the aversive nature of conflicts.
However, and for the sake of completeness, it should be noted that there exist at least two studies that were not able to find any evidence in favor of the affective valence of conflict stimuli. Schacht and colleagues (Schacht, Dimigen, & Sommer, 2010; Schacht, Nigbur, & Sommer, 2009) registered physiological markers of arousal and emotion in a go/no-go paradigm and found reduced startle blink and skin conductance responses in (conflict) no-go trials, as compared with (nonconflict) go trials, denoting, if anything, rather reduced arousal and less negative valence for no-go trials. However, go and no-go trials differ not only with respect to their (presumed) affective valence, but also, obviously so, with respect to other processing characteristics (like motor affordances). In contrast, Renaud and Blondin (1997) let participants perform a variation of the Stroop task with equal motor affordances for high- and low-conflict stimuli while recording their heart rates. More precisely, conflict was manipulated block-wise: Participants performed blocks of either only incongruent or only neutral (colored XXX) trials. The authors found increased heart rates in the conflict (incongruent stimuli), as compared with nonconflict (neutral stimuli), blocks. In the same line, Kobayashi, Yoshino, Takahashi, and Nomura (2007) reported enhanced skin conductance responses (SCRs) associated with incongruent trials in a Stroop task. Taken together, the findings of Renaud and Blondin and Kobayashi et al. can be taken as evidence for the increased effort associated with conflict stimuli. Higher effort, in turn, is considered to be experienced as aversive (Song & Schwarz, 2008). This is also in line with Hajcak, McDonald, and Simons (2004), who reported an association of enhanced heart rate and SCRs with negative affect in response to error conduction. In sum, there is now physiological, as well as psychological, evidence for the aversive nature of conflicts.
One limitation of the present study rests on the working principle of the affective priming paradigm. In this paradigm, participants do not have to react overtly to the primes. Since we used conflict and nonconflict Stroop primes as affective primes in our study, this raises the question of whether passive viewing of conflict primes indeed induced (response) conflict in the present paradigm. In fact, in the original conflict model as proposed by Botvinick and colleagues, conflict was measured over the response (i.e., output) layer (Botvinick et al., 2001), suggesting that response conflict is actually a mandatory precondition for control adjustments to occur. However, the authors also admitted that, in principle, such conflict could occur at any other representational level. More precisely, they stated that conflict might be triggered by any “simultaneous activation of incompatible representations” (p. 680). Furthermore, van Veen et al. (2004) pointed out that conflict monitoring has repeatedly been found not to be restricted to the response level. For example, there is evidence for ACC activation for response as well as perceptual and semantic conflict in a global/local task (Weissman et al., 2003) and in the nonresponse condition of a Stroop task (Milham, Banich, & Barad, 2003). And finally, in our study, it can be assumed that at least preparatory motor processes were present in both experiments. Note that participants had to monitor for events that required motor reactions to Stroop primes (i.e., catch trials). These events were denoted by both the word and color dimension of the primes; that is, together with the finding of a very low number of catch trial omissions, it is very likely that the Stroop primes induced not only stimulus, but probably also response conflict. As a side note, the functional boundaries of response and stimulus conflict are still not drawn clearly in the literature; Notebaert and Verguts (2006), for example, found enhanced conflict adaptation effects for stimulus conflict, but not for response conflict.
The notion of the ACC as a detector of both cognitive and affective conflict has been proposed earlier (Bush, Luu, & Posner, 2000). Interestingly, while Bush et al. ascribed the cognitive and affective monitoring functions to separate regions of the ACC (dorsal and ventral ACC, respectively), Shackman et al. (2011) argued against this segregated view of ACC function in a recent review. Indeed, the authors presented strong evidence for conjoint anterior midcingulate cortex (mACC) activation by cognitive control, negative affect, and pain. This fits perfectly with the idea that this region of the ACC serves the function of an aversive signal detector, as has been suggested by Botvinick (2007), whose integrative account of ACC function was the starting point for our investigation. The author proposed that two long-standing theories of ACC function—namely, the conflict monitoring theory (Botvinick et al., 2001) and the outcome evaluation account (Holroyd & Coles, 2002)—might converge on the detection of aversive signals in general. The present results strongly support the notion of the ACC as a detector and indicator of aversive signals. However, given that our conclusions are based entirely on behavioral data, it will be vital to investigate the neural mechanisms underlying the conflict priming effect in future studies. More precisely, a consequential assumption that could be tested using functional MRI is whether and how valence judgments of targets actually covary with the strength of prime-induced ACC activation. Furthermore, assuming that the ACC only detects the aversive signal conveyed by a conflict, future studies should elucidate the role of brain regions other than the ACC that are involved in the further processing and generation of the negative judgments after the (aversive) conflict is detected. For instance, it is conceivable that functional connectivity between the ACC and affect-related regions (e.g., amygdala, striatum, or ventromedial prefrontal cortex) varies as a function of conflict priming.
To conclude, the results of the conflict priming paradigm presented here, together with previous more or less direct evidence, show that conflict indeed is registered as an aversive signal (Dreisbach & Fischer, 2012a; Schouppe et al., 2012; van Steenbergen et al., 2009, 2010). Given the further evidence that aversive stimuli have been shown to induce sequential processing adjustments even in the absence of any response conflict (Dreisbach & Fischer, 2011), it can be assumed that the conflict monitoring theory might actually be applicable to any aversive signal in the ongoing processing stream. Our results thus add to the growing literature showing that affective signals play an important role in sequential action control (see Dreisbach & Fischer, 2012b, for a review).
Notes
Online English–Chinese dictionary available at http://www.mdbg.net/chindict/chindict.php.
In Experiment 1a, as in the previous study by Dreisbach and Fischer (2012a), the experiment started with a short block of 24 Stroop stimuli, where participants had to name the color of the words to make themselves familiar with the (aversive character) of the Stroop primes. However, because the error rate for catch trials in Experiment 1a was very low, the short Stroop block was dropped in Experiment 1b, since catch trials successfully ensured the processing of the Stroop primes.
For the sake of completeness, mean RTs for each prime–target combination were entered into a 2 (prime congruence: conflict/nonconflict) × 2 (judgment: positive/negative) ANOVA with repeated measures. No main effect or interaction reached significance (all Fs < 2.3, all ps > 1.3). For mean RTs, see the Appendix, Table 1.
References
Botvinick, M. W. (2007). Conflict monitoring and decision making: Reconciling two perspectives on anterior cingulated function. Cognitive, Affective, & Behavioral Neuroscience, 7, 356–366.
Botvinick, M. M., Braver, T. S., Barch, D. M., Carter, C. S., & Cohen, J. D. (2001). Conflict monitoring and cognitive control. Psychological Review, 108, 624–652.
Botvinick, M. M., Nystrom, L. E., Fissell, K., Carter, C. S., & Cohen, J. D. (1999). Conflict monitoring versus selection-for-action in anterior cingulate cortex. Nature, 402, 179–181.
Braem, S., Verguts, T., Roggeman, C., & Notebaert, N. (2012). Reward modulates adaptations to conflict. Cognition, 125(2), 324–332.
Bush, G., Luu, P., & Posner, M. I. (2000). Cognitive and emotional influences in anterior cingulate cortex. Trends in Cognitive Sciences, 4, 215–222.
Casasanto, D. (2009). Embodiment of abstract concepts: Good and bad in right- and left- handers. Journal of Experimental Psychology. General, 138(3), 351–367.
Chang, B., & Mitchell, C. J. (2009). Processing fluency as a source of salience asymmetries in the Implicit Association Test. Quarterly Journal of Experimental Psychology, 26, 1–25.
Dreisbach, G., & Fischer, R. (2011). If it's hard to read… try harder! Processing fluency as signal for effort adjustments. Psychological Research, 75, 376–383.
Dreisbach, G., & Fischer, R. (2012a). Conflicts as aversive signals. Brain and Cognition, 72, 94–98.
Dreisbach, G., & Fischer, R. (2012b). The role of affect and reward in the conflict-triggered adjustment of cognitive control. Frontiers in Human Neuroscience, 6:342. doi:10.3389/fnhum.2012.00342
Egner, T. (2007). Congruency sequence effects and cognitive control. Cognitive, Affective, & Behavioral Neuroscience, 7, 380–390.
Eisenberger, N. I., Lieberman, M. D., & Williams, K. D. (2003). Does rejection hurt? An fMRI study of social exclusion. Science, 302, 290–292.
Fazio, R. H. (2001). On the automatic activation of associated evaluations: An overview. Cognition and Emotion, 15(2), 115–141.
Fazio, R. H., Sanbonmatsu, D. M., Powell, M. C., & Kardes, F. R. (1986). On the automatic activation of attitudes. Journal of Personality and Social Psychology, 50(2), 229–238.
Gratton, G., Coles, M. G. H., & Donchin, E. (1992). Optimizing the use of information: Strategic control of activation of responses. Journal of Experimental Psychology. General, 121, 480–506.
Greenwald, A. G., McGhee, D. E., & Schwartz, J. L. K. (1998). Measuring individual differences in implicit cognition: The Implicit Association Test. Journal of Personality and Social Psychology, 74, 1464–1480.
Gulan, T., & Valerjev, P. (2010). Semantic and related types of priming as a context in word recognition. Review of Psychology, 17(1), 53–58.
Hajcak, G., McDonald, N., & Simons, R. F. (2004). Error-related psychophysiology and negative affect. Brain and Cognition, 56, 189–197.
Hanslmayr, S., Pastötter, B., Bäuml, K.-H., Gruber, S., Wimber, M., & Klimesch, W. (2008). The electrophysiological dynamics of interference during the Stroop task. Journal of Cognitive Neuroscience, 20(2), 215–225.
Holroyd, C. B., & Coles, M. G. H. (2002). The neural basis of human error processing: Reinforcement learning, dopamine, and the error related negativity. Psychological Review, 109, 679–709.
Kerns, J. G., Cohen, J. D., MacDonald, A. W., III, Cho, R. Y., Stenger, V. A., & Carter, C. S. (2004). Anterior cingulate conflict monitoring and adjustments in control. Science, 303, 1023–1026.
Kobayashi, N., Yoshino, A., Takahashi, Y., & Nomura, S. (2007). Autonomic arousal in cognitive conflict resolution. Autonomic Neuroscience - Basic, 132, 70–75.
Kornblum, S., Hasbroucq, T., & Osman, A. (1990). Dimensional overlap: Cognitive basis for stimulus–response compatibility–a model and taxonomy. Psychological Review, 97(2), 253–270.
Liotti, M., Woldorff, M. G., Perez, R., & Mayberg, H. S. (2000). An ERP study of the temporal course of the Stroop color-word interference effect. Neuropsychologia, 38(5), 701–711.
MacDonald, A. W., Cohen, J. D., Stenger, V. A., & Carter, C. S. (2000). Dissociating the role of the DLPFC & ACC in cognitive control. Science, 288, 1835–1838.
Milham, M. P., Banich, M. T., & Barad, V. (2003). Competition for priority in processing increases prefrontal cortex's involvement in top-down control: An event-related fMRI study of the Stroop Task. Cognitive Brain Research, 17, 212–222.
Miller, E. K., & Cohen, J. D. (2001). Integrative theory of PFC function. Annual Review of Neuroscience, 24, 167–202.
Murphy, S. T., Monahan, J. L., & Zajonc, R. B. (1995). Additivity of nonconscious affect: Combined effects of priming and exposure. Journal of Personality and Social Psychology, 69, 589–602.
Murphy, S. T., & Zajonc, R. B. (1993). Affect, cognition, and awareness: Affective priming with suboptimal and optimal stimulus. Journal of Personality and Social Psychology, 64, 723–739.
Nieuwenhuis, S., Yeung, N., Holroyd, C. B., Schurger, A., & Cohen, J. D. (2004). Sensitivity of electrophysiological activity from medial frontal cortex to utilitarian and performance feedback. Cerebral Cortex, 14, 741–747.
Notebaert, W., Soetens, E., & Melis, A. (2001). Sequential analysis of a Simon task. Evidence for an attention-shift account. Psychological Research, 65, 170–184.
Notebaert, W., & Verguts, T. (2006). Stimulus conflict predicts conflict adaptation in a numerical flanker task. Psychonomic Bulletin & Review, 13(6), 1078–1084.
Payne, B. K., Cheng, C. M., Govorun, O., & Stewart, B. (2005). An inkblot for attitudes: Affect misattribution as implicit measurement. Journal of Personality and Social Psychology, 89, 277–293.
Payne, B. K., Shimizu, Y., & Jacoby, L. L. (2005). Mental control and visual illusions: Toward explaining race-biased weapon identifications. Journal of Experimental Social Psychology, 41, 36–47.
Rainville, P. (2002). Brain mechanisms of pain affect and pain modulation. Current Opinion in Neurobiology, 12, 195–204.
Reber, R., Schwarz, N., & Winkielman, P. (2004). Processing fluency and aesthetic pleasure: Is beauty in the perceiver’s processing experience? Personality and Social Psychology Review: An Official Journal of the Society for Personality and Social Psychology, Inc, 8(4), 364–382.
Renaud, P., & Blondin, J.-P. (1997). The stress of Stroop performance: Physiological and emotional responses to color-word interference, task pacing, and pacing speed. International Journal of Psychophysiology, 27, 87–97.
Schacht, A., Dimigen, O., & Sommer, W. (2010). Emotions in Cognitive Conflicts are Not Aversive but are Task Specific. Cognitive, Affective, and Behavioral Neursocience, 10, 349–356.
Schacht, A., Nigbur, R., & Sommer, W. (2009). Emotions in Go/ NoGo conflicts. Psychological Research, 73, 843–856.
Schouppe, N., De Houwer, J., Ridderinkhof, R., & Notebaert, W. (2012). Conflict: Run! Reduced stroop interference with avoidance responses. Quarterly Journal of Experimental Psychology, 65(6), 1052–1058.
Scott, G. G., O’Donnell, P. J., Leuthold, H., & Sereno, S. C. (2009). Early emotion word processing: Evidence from event-related potentials. Biological Psychology, 80, 95–104.
Shackman, A. J., Salomons, T. V., Slagter, H. A., Fox, A. S., Winter, J. J., & Davidson, R. J. (2011). The integration of negative affect, pain and cognitive control in the cingulate cortex. Nature Reviews Neuroscience, 12, 154–167.
Singer, T., Seymour, B., O’Doherty, J., Kaube, H., Dolan, R. J., & Frith, C. D. (2004). Empathy for pain involves affective but not sensory components of pain. Science, 303, 1157–1162.
Song, H., & Schwarz, N. (2008). If it’s hard to read, it’s hard to do: Processing fluency affects effort prediction and motivation. Psychological Science, 19, 986-988.
Stürmer, B., Leuthold, H., Soetens, E., Schröter, H., & Sommer, W. (2002). Control over location-based response activation in the Simon task: Behavioral and electrophysiological evidence. Journal of Experimental Psychology. Human Perception and Performance, 28, 1345–1363.
Unkelbach, C., Fiedler, K., Bayer, M., Stegmüller, M., & Danner, D. (2008). Why positive information is processed faster: The density hypothesis. Journal of Personality and Social Psychology, 95(1), 36–49.
van Steenbergen, H., Band, G. P. H., & Hommel, B. (2009). Reward counteracts conflict adaptation. Evidence for a role of affect in executive control. Psychological Science, 20(12), 1473–1477.
van Steenbergen, H., Band, G. P. H., & Hommel, B. (2010). In the mood for adaptation: How affect regulates conflict-driven control. Psychological Science, 21, 1629–1634.
Van Veen, V., Holroyd, C. B., Cohen, J. D., Stenger, V. A., & Carter, C. S. (2004). Errors without conflict: Implication for performance monitoring theories of anterior cingulate cortex. Brain and Cognition, 56, 267–276.
Võ, M. L.-H., Conrad, M., Kuchinke, L., Urton, K., Hofmann, M. J., & Jacobs, A. M. (2009). The Berlin Affective Word List Reloaded (BAWL-R). Behavior Research Methods, 41(2), 534–538.
Weissman, D. H., Giesbrecht, B., Song, A. W., Mangun, G. R., & Woldorff, M. G. (2003). Conflict monitoring in the human anterior cingulate cortex during selective attention to global and local object features. NeuroImage, 19, 1361–1368.
Winkielman, P., & Cacioppo, J. T. (2001). Mind at ease puts a smile on the face: Psychophysiological evidence that processing facilitation elicits positive affect. Journal of Personality and Social Psychology, 81(6), 989–1000.
Wühr, P., & Ansorge, U. (2005). Exploring trial-by-trial modulations of the Simon effect. The Quarterly Journal of Experimental Psychology. A, 58(4), 705–731.
Yeung, N., Botvinick, M. M., & Cohen, J. D. (2004). The Neural Basis of Error Detection: Conflict Monitoring and the Error-Related Negativity. Psychological Review, 111(4), 931–959.
Zajonc, R. B. (1968). Attitudinal effects of mere exposure. Journal of Personality and Social Psychology, 9, 1–27.
Author Notes
This research was supported by a grant to the second author from Deutsche Forschungsgemeinschaft (DFG: DR 392/6-1).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Fritz, J., Dreisbach, G. Conflicts as aversive signals: Conflict priming increases negative judgments for neutral stimuli. Cogn Affect Behav Neurosci 13, 311–317 (2013). https://doi.org/10.3758/s13415-012-0147-1
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13415-012-0147-1