
Abstract
Although it is often assumed that a physically salient stimulus automatically captures attention even when it is irrelevant to a current task, the signal-suppression hypothesis proposes that observers can actively suppress a salient-but-irrelevant distractor. However, it is still unknown whether suppression alone (i.e., without target enhancement) is potent enough to override attentional capture by a salient singleton in an otherwise-homogeneous background. The current study addressed this issue. On search trials (70% of trials), participants searched for a shape target on trials that either did or did not contain an irrelevant color singleton. The effects of learning to suppress the color of the singleton were examined on interleaved probe trials (30% of trials). On these trials, participants searched for a probe target letter; those letters were presented on four ovals (one colored oval and three gray ovals). Each colored oval was a singleton that was one of three types: the color of the distractor on search trials, the color of the target on search trials, or a neutral color that had not appeared on search trials. Responses were faster for the probe target on a neutral-colored or target-colored item than on a gray-colored item; however, responses were slower for the probe target on a distractor-colored item than on a gray-colored item. The results demonstrate a powerful suppression mechanism overriding attentional capture by a singleton item.
Similar content being viewed by others


Introduction
It is often assumed that physically salient items that differ from other stimuli in terms of a basic feature (i.e., feature singletons such as a red item among green items) automatically attract our visual attention. According to stimulus-driven theories, physically salient items automatically capture visual attention regardless of an observer’s current goals and intentions (e.g., Theeuwes, 1992). In a classic paradigm developed by Theeuwes (1992), when participants searched for a circle target among diamonds to report the orientation of a line inside the circle, responses to the target were slowed on trials on which a singleton color distractor appeared. This singleton-presence cost demonstrates attentional capture by the color singleton, slowing attentional allocation to the target. Such a bottom-up process is assumed to determine attentional priority (Itti & Koch, 2000). In some models it interacts with a top-down process in which attention is guided by voluntary goals and intentions (Folk, Remington, & Johnston, 1992) as well as the history of attentional selection (Awh, Belopolsky, & Theeuwes, 2012; Theeuwes, 2018, 2019).
Recently, the signal-suppression hypothesis has been proposed, suggesting that salient stimuli automatically generate an attentional priority signal but that signal can be actively suppressed by an inhibitory control process before attention is captured (Sawaki & Luck, 2010). The original evidence for active suppression came from event-related potential (ERP) studies focusing on the Pd component, which is often considered to be inhibition-related (Hickey, Di Lollo, & McDonald, 2009). Subsequent studies provided behavioral data supporting the signal-suppression hypothesis (Gaspelin, Leonard, & Luck, 2015; Gaspelin & Luck, 2018b; see Wyble et al., 2020, for a model-based simulation of the data of Gaspelin et al., 2015). Those behavioral studies used a capture-probe paradigm to test suppression of salient singletons. On 70% of trials (search trials), participants searched for a shape-defined target (e.g., a diamond) among four different shapes and reported whether the target had a dot on the left or right side. Half of the search trials were singleton-present trials containing a color singleton distractor while the other half of the search trials were singleton-absent trials without a color singleton distractor. Search trials revealed a search benefit on singleton-present trials; responses were faster when a color singleton was present.Footnote 1 On the other 30% of trials (probe trials), alphabet letters were briefly presented on the shapes, and participants were asked to report as many letters as possible. The critical finding was that on probe trials, the letter on a singleton distractor was less likely to be reported compared to those on non-singleton distractors or a target (see also Feldmann-Wüstefeld, Busch, & Schubö, 2020, for additional evidence, both behavioral and electrophysiological, of below-baseline suppression of salient distractors). Although these results were taken as evidence that salient singleton distractors can be actively suppressed, the results do not necessarily support a strong form of the signal-suppression hypothesis because the effects found in their study may not be solely attributable to distractor-feature suppression; they may also be explained by target-feature enhancement. Since a salient color singleton was presented among other items that consisted of a target and several other items that also had the color of the target, the same results can also be explained by assuming that participants boosted the target color value. If the results arise from target-feature enhancement rather than distractor-feature suppression or from a combination of both, it weakens the argument that a salient singleton is actively suppressed.
In an attempt to address this issue, Chang and Egeth (2019) used a variant of the capture-probe paradigm and showed that distractor-feature suppression was independent of target-feature enhancement. In Chang and Egeth’s (2019) study, on 70% of trials (search trials), participants indicated whether a black dot on a diamond target was on its left or right side. Those search trials were either singleton-present or singleton-absent trials. Participants responded more quickly on singleton-present than on singleton-absent trials, showing a singleton-presence benefit. On the other 30% of trials (probe trials), participants were presented with four ovals, each with a letter inside of it, and were asked to report which of two probe target letters (A or B) was present. Each oval was in a different color; one oval was either the color of the target or the color of the distractor from search trials, and the other three were neutral (i.e., three colors that were not used on search trials). Faster and more accurate responses were made when the probe target appeared on a target-colored item than when it appeared on a neutral-colored item, whereas slower and less accurate responses were made when the probe target appeared on a distractor-colored item than when it appeared on a neutral-colored item. The results demonstrated that a distractor feature can be suppressed independently of enhancement of a target feature, and that both enhancement and suppression contribute to guiding attention. This finding suggests that the previous results may reflect a combined effect of distractor-feature suppression and target-feature enhancement. Although Chang and Egeth (2019) tested each mechanism separately, since target-feature enhancement and distractor-feature suppression were tested with heterogeneous displays, the results cannot tell us whether suppression alone is strong enough to override capture by a salient singleton.
To our knowledge, none of the previous studies provide direct evidence for suppression overriding attentional capture by salient singletons; there have been no demonstrations that the strongest stimulus in a scene can be actively suppressed. Gaspelin and colleagues only showed suppression of a singleton distractor when that singleton was presented among target-colored items (Gaspelin, Gaspar, & Luck, 2019; Gaspelin et al., 2015; Gaspelin, Leonard, & Luck, 2017; Gaspelin & Luck, 2018b), while Chang and Egeth (2019) only showed suppression of a distractor-colored item among heterogeneous neutral-colored items. The current research sought to determine whether suppression alone (i.e., without target enhancement) is potent enough to override attentional capture by a salient singleton in an otherwise-homogeneous background. To maximize salience of the singleton distractor, we used a new probe display where a color singleton oval was presented among three gray ovals. All items were the same shape, and the singleton was a color while other items were all the same shade of gray. This new probe display was expected to maximize pop-out (Duncan & Humphrey, 1989; Nothdurft, 1993), and it allows a rigorous test of whether a strong salience signal can be suppressed.
Method
Participants
Sixty undergraduate students (mean age = 19.0 years; 21 male, 39 female) at Johns Hopkins University participated. We used the same sample size (N = 60) as a previous study that used a paradigm analogous to the one used in the present study (Chang & Egeth, 2019). Data collection stopped once we reached 60 participants. All participants had normal or corrected-to-normal vision based on self-report. Informed consent was obtained from all participants, and the protocols were approved by the Johns Hopkins Homewood Institutional Review Board.
Apparatus
The experiment was conducted on a Dell Precision T3400 2.33-GHz computer. Stimulus presentation was performed using programs written in MATLAB (The MathWorks, Natick, MA, USA) and Psychophysics Toolbox software (Brainard, 1997). Stimuli were presented on a Dell 1708FP monitor (refresh rate = 60 Hz, resolution = 1,280 × 1,024 pixels) and viewed at a distance of approximately 60 cm from the participants. Responses were made by pressing the “left” or “right” button of a game controller with the left and right index fingers.
Stimuli, design, and procedure
All participants were provided with written and oral descriptions of the stimuli and procedures. Participants were instructed to maintain fixation on the centrally presented fixation cross during the entire trial. Search and probe trials were randomly intermixed.
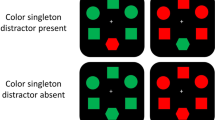
Both search and probe trials contained a fixation point, a search array, and a feedback display. All stimuli were presented on a black background. The search and probe displays were arranged to form a diamond pattern (5.17° in width and height, from shape center to shape center, see Fig. 1). Three colors (red [CIE x = 0.640, y = 0.330], green [x = 0.300, y = 0.600], blue [x = 0.150, y = 0.060]) and one gray (CIE x = 0.313, y = 0.329) were used.

Example displays from the trial types used in the experiment. On search trials, participants made a speeded response to the location of a dot (left or right) inside the search target (diamond). On probe trials, participants made a speeded choice response about the presence of a probe target (A or B). Probe targets appeared equally often at each location. Critical colors were fully counterbalanced across participants in the experiment, but in this example, the neutral color is blue, the target color is green, and the distractor color is red
On search trials (70% of trials), four different shapes were presented, and participants were asked to report whether a black dot was on the left or right side of the target shape by pressing a left or right button as quickly and accurately as possible. The search target was always the diamond. Search displays contained one diamond (1.68° × 1.68°), one circle (1.53° × 1.53°), one square (1.53° × 1.53°), and one hexagon (1.53° × 1.53°). Each shape contained a 0.15° black dot located 0.3° from either the left or right side of the shape. Target location and dot side varied randomly. On 50% of these trials, all items were colored homogeneously (i.e., singleton-absent), and on the other half of trials, one item was drawn in a different color (i.e., singleton-present). The location of this color singleton distractor was random except that it was never the target location.
Search trials began with the presentation of a blank screen for 500 ms, followed by a fixation screen for 1,000 ms. Subsequently, the search array appeared for 2,000 ms. If participants did not respond by 2,000 ms, a timeout display appeared displaying the text “Too Slow” for 500 ms. A feedback display followed a response for 500 ms. The feedback display contained the word “Correct!” for correct trials or “Incorrect!” for incorrect trials in the middle of the screen.
On probe trials (30% of trials), four ovals (a shape that was never presented on search trials) were presented, and a letter was presented inside each item. One of the letters was either “A” or “B,” and the task was to report which probe target letter (A or B) was present by pressing a left or right button as quickly and accurately as possible. Probe displays contained four identical oval shapes (1.53° × 1.21°). Upper-case letters (0.75° high) were presented in white at the center of each oval using Arial font. Probe targets appeared equally often at each of the four locations. The other three letters were selected randomly without replacement from the English alphabet except for “I” and “O.” The location of the probe target was varied randomly.
On probe trials, three of the four ovals were in a gray color, and one of the ovals was in a critical color. There were three possibilities for this critical color, and they defined a crucial experimental manipulation. In one case, the critical color was the color that had never been presented on search trials (i.e., neutral colorFootnote 2; blue in the example shown in Fig. 1). This neutral-color condition serves as a baseline for the remaining two conditions. In another case, the critical color was the color in which the target shape appeared on search trials (green in the example of Fig. 1). In the third case, the critical color was the color that served as the singleton distractor color on search trials (red in the example shown in Fig. 1). We will refer to these three kinds of probe trials as neutral-colored singleton, target-colored singleton, and distractor-colored singleton trials, respectively, but keep in mind that these labels are derived from the status of these colors on search trials, not on probe trials. The probe target appeared at each location equally often; it appeared on a singleton item (i.e., neutral-colored, target-colored, or distractor-colored item) on 25% of trials and on a random gray-colored item on 75% of trials.
Probe trials began with the presentation of a blank screen for 500 ms followed by a fixation screen for 1,000 ms. Subsequently, the probe array appeared for 100 ms. If participants did not respond by 100 ms, the probe array disappeared and a blank screen appeared until response. A feedback display was presented for 500 ms following responses. The feedback display contained the word “Correct!” for correct trials or “Incorrect!” for incorrect trials in the middle of the screen.
A set of three colors (red, green, and blue) was used for the critical colors, and their roles (neutral, target, and distractor colors) in the experiment were fully counterbalanced across participants, eliminating any confounding effects of colors. For a given participant, the role of a particular color was constant for the duration of the experiment. One-third of probe trials were neutral-colored singleton trials, one-third of probe trials were target-colored singleton trials, and the other one-third of probe trials were distractor-colored singleton trials. In all cases, the critical-colored items were a color singleton among gray items.
The experiment began with 48 practice trials for the probe task but all the items were gray on these practice trials. Following the practice trials of the probe task, participants performed 48 trials of practice with the search task, and this task was the same as the one in the main experiment. Practice trials were excluded from the analyses reported below. The main experiment consisted of 336 search trials and 144 probe trials, resulting in 480 trials in total.
Results
Search-task
Mean accuracy was 96.8% and 96.7% for singleton-present and singleton-absent trials, respectively (no significant difference; t(59) = .381, p = .705, d = 0.049). All reaction times faster than 100 ms as well as latencies from incorrect trials were excluded from the analysis on an a priori basis. Based on a modified recursive trimming procedure (Van Selst & Jolicoeur, 1994), reaction times 3.5 standard deviations above or below the mean for each participant were subsequently removed. Altogether, this resulted in the elimination of 3% of all search trials.
A t-test was conducted on singleton presence for singleton-present and singleton-absent trials. Responses on search trials were faster when the color singleton was present (M = 679 ms) than when it was absent (M = 686 ms); there was a 7-ms singleton-presence benefit, t(59) = 2.593, p = .012, d = 0.335.
Probe-task
Incorrect trials and outlier latencies were excluded as in the search task except that reaction times slower than 5,000 ms (0.08% of probe trials) were also excluded from the analysis prior to the trimming procedure. Trials were also excluded from the analysis of probe data if two or more consecutive preceding trials were probe trials, because consecutive probing could temporarily disrupt the established attentional set for the target and singleton distractor, and distort probe responses (Gaspelin & Luck, 2018a). Altogether, these exclusions resulted in the elimination of 8% of probe trials. Response time and accuracy (see Table 1) were both analyzed in 3 × 2 repeated-measures analyses of variance (ANOVA). One factor was critical color; whether the neutral-colored, target-colored, or distractor-colored singleton was present on a probe trial. The other factor was the location of a probe target (A or B) on a probe trial; this was either on a critical-colored item or on one of the gray-colored items. All subsequent multiple comparisons were Bonferroni corrected.
Reaction time
The main effect of critical color was significant, F(2, 118) = 6.583, p = .002, \( {\eta}_p^2 \) = .100. Pairwise comparisons revealed that responses were slower on distractor-colored singleton trials (M = 741 ms) than on both target-colored singleton trials (M = 723 ms), t(59) = 3.24, p = .006, d = 0.419, and neutral-colored singleton trials (M = 719 ms), t(59) = 2.69, p = .028, d = 0.347. The main effect of probe-target location was also significant, F(1, 59) = 26.929, p < .001, \( {\eta}_p^2 \) = .313. Responses were faster when a probe target was on critical-colored items (M = 714 ms) than on gray-colored items (M = 742 ms). More importantly, the interaction between critical color and probe-target location was significant, F(2, 118) = 25.891, p < .001, \( {\eta}_p^2 \) = .305. Pairwise comparisons revealed that on neutral-colored singleton trials responses were faster when the probe target was presented on the neutral-colored item (M = 701 ms) than on one of the gray items (M = 738 ms), t(59) = 3.62, p = .001, d = 0.468, showing a 37-ms singleton-facilitation effect. On target-colored singleton trials responses were faster when the probe target was presented on the target-colored item (M = 690 ms) than on one of the gray items (M = 755 ms), t(59) = 8.75, p < .001, d = 1.130, showing a 65-ms singleton-facilitation effect. In contrast, and critically, on distractor-colored singleton trials responses were slower when the probe target was presented on the distractor-colored item (M = 750 ms) than on one of the gray items (M = 731 ms), t(59) = 2.32, p = .024, d = 0.299, showing a 19-ms singleton-inhibition effect.
To look at these results through a slightly different lens, for each critical color condition, singleton facilitation scores were calculated (reaction time to a probe target on a gray-colored item – reaction time to a probe target on a critical-colored item). A repeated-measures one-way ANOVA was conducted to test the effect of critical color on singleton facilitation scores. The effect of critical color was significant, F(2, 118) = 25.891, p < .001, \( {\eta}_p^2 \) = .305 (see Fig. 2a). Pairwise comparisons revealed that the singleton effect on distractor-colored singleton trials (M = -19 ms) was significantly different from those on target-colored singleton trials (M = 65 ms), t(59) = 7.63, p < .001, d = 0.985, and neutral-colored singleton trials (M = 37 ms), t(59) = 4.51, p < .001, d = 0.582. However, the difference in the singleton effects on target-colored singleton and neutral-colored singleton trials did not reach significance, p = .077.

Singleton facilitation scores for (a) reaction time (RT) and (b) error rate as a function of singleton color. Singleton facilitation scores were calculated by subtracting RT and error rate for a probe target on a singleton from RT and error rate for a probe target on a gray item. Neutral color refers to the color that had never been presented on search trials, and target and distractor colors refer to the roles those colors had on search trials. Error bars represent standard errors of the mean, corrected for within-subjects designs (Cousineau, 2005)
Error rate
The results from error data conform to the results from RT data. The main effect of critical color was significant, F(2, 118) = 9.343, p < . 001, \( {\eta}_p^2 \) = .137. Pairwise comparisons revealed that error rate was higher on distractor-colored singleton trials (M = 9.9%) than on target-colored singleton trials (M = 6.4%), t(59) = 4.38, p < .001, d = 0.565, and on neutral-colored singleton trials (M = 6.7%), t(59) = 3.20, p = .008, d = 0.413. The main effect of probe-target location was also significant, F(1, 59) = 5.104, p = .028, \( {\eta}_p^2 \) = .08. Error rate was lower when the probe target was on a critical colored item (M = 6.9%) than when it was on gray-colored items (M = 8.4%). More importantly, the interaction between critical color and probe-target location was significant, F(2, 118) = 26.677, p < .001, \( {\eta}_p^2 \) = .311. Pairwise comparisons revealed that on neutral-colored singleton trials error rate was lower when the probe target was presented on the neutral-colored item (M = 4.4%) than on one of the gray items (M = 9.0%), t(59) = 4.27, p < .001, d = 0.552. On target-colored singleton trials error rate was lower when the probe target was presented on the target-colored item (M = 3.8%) than on one of the gray items (M = 9.0%), t(59) = 5.10, p < .001, d = 0.658. On the other hand, on distractor-colored singleton trials error rate was higher when the probe target was presented on the distractor-colored item (M = 12.6%) than on one of the gray items (M = 7.2%), t(59) = 4.15, p < .001, d = 0.536.
As in the RT analysis, to further examine these results, for each critical color condition, singleton facilitation scores were calculated (error rate for a probe target on a gray-colored item – error rate for a probe target on a critical-colored item). A repeated-measures one-way ANOVA was conducted to test the effect of critical color on singleton facilitation scores. The effect of critical color was significant, F(2, 118) = 26.677, p < .001, \( {\eta}_p^2 \) = .311 (see Fig. 2b). Pairwise comparisons revealed that the singleton effect on distractor-colored singleton trials (M = -5.4%) was significantly different from those on target-colored singleton trials (M = 5.1%), t(59) = 6.56, p < .001, d = 0.847, and neutral-colored singleton trials (M = 4.7%), t(59) = 5.32, p < .001, d = 0.686. However, the difference in the singleton effects on target-colored singleton and neutral-colored singleton trials was not significantly different, p = 1.
Discussion
The signal-suppression hypothesis argues that a salience signal generated by a singleton can be actively suppressed before that stimulus captures attention. Even though previous studies provided evidence supporting the signal-suppression hypothesis (e.g., Gaspelin et al., 2015), it was still unknown whether suppression alone (i.e., without target enhancement) is sufficiently potent to effect such suppression for a highly salient stimulus in an otherwise-homogeneous background. In the present study, we directly tested whether salient stimuli can be suppressed.
On search trials, participants learned the features (colors) of a target and a distractor with consistent color roles, and showed a singleton presence benefit. On probe trials, when a color singleton was presented among homogeneous gray items, rendering the color singleton as a hard-to-ignore singleton, a distractor-colored singleton was suppressed in comparison with gray items while a target-colored singleton and a neutral-colored singleton were facilitated in comparison with the same gray items.
On probe trials, all the items were in the same shape (oval), and one of the four ovals had a color (red, green, or blue) among the other gray ovals. Such displays were used to maximize the possibility of attentional capture by a color singleton. Indeed, we found a 37-ms singleton facilitation effectFootnote 3 on neutral-colored singleton trials; a probe target was identified faster when it was on a neutral-colored singleton item compared to when it was on a gray-colored item. Since the neutral color never appeared on search trials, it functions as a baseline for singleton-related effects. Target-colored singleton trials showed a 65-ms singleton facilitation effect; a probe target was identified faster when it was on a target-colored singleton item compared to when it was on a gray-colored item. The larger facilitation effect on target-colored singleton trials (65 ms) than that on neutral-colored singleton trials (37 ms) seems to reflect the additive effect of target-feature enhancement and the singleton-facilitation effect, but this suggestion is offered tentatively as the difference was not statistically significant. More interestingly, a 19-ms singleton-inhibition effect was found on distractor-colored singleton trials; a probe target was identified more slowly when it was on a distractor-colored singleton item compared to when it was on a gray-colored item. The distractor feature was suppressed even when it was a highly salient color singleton among homogeneous gray items; distractor-feature suppression overrode attentional capture by a salient singleton, which would otherwise have shown a singleton facilitation effect.
Some might argue that the singleton inhibition effect may not reflect pure suppression without target enhancement if one assumes that participants enhance large swaths of the color space. In other words, participants may have enhanced colors other than the singleton color, and that could produce the same pattern of results as observed on distractor-colored singleton trials. It is difficult to falsify this sort of upweighting-only model without further constraints. However, it is worth noting that Chang and Egeth (2019) addressed this issue in discussing the results from a similar experiment. They showed that a specific color (i.e., the target color from the search task) was enhanced among heterogenous neutral colors on their probe trials, and furthermore the effect was not modulated by target-distractor similarity. This demonstrates that observers enhance a specific color rather than large swaths of the color space including the target color. The present finding that a larger singleton-facilitation effect was observed on target-colored singleton trials than on neutral-colored singleton trials is consistent with the previous finding (Chang & Egeth, 2019). Combining the previous and the current findings, it seems reasonable to conclude that suppression alone is potent enough to override attentional capture by salient singletons, although further research is clearly called for.
The current results are consistent with a model of suppression that proposes suppression occurs on the basis of specific feature values (e.g., redness; Gaspelin & Luck, 2018b). Gaspelin and Luck (2018b) examined three possible mechanisms of visual suppression (first-order feature suppression, second-order feature suppression, and global-salience suppression), and showed that participants learn to suppress the singleton color, not color singletons per se or general salience. In the current study, each critical color was presented as a singleton among gray items on probe trials. If participants were learning to suppress any color singleton or overall salience, the target-colored and neutral-colored singletons should have been suppressed. However, it was not the case; the distractor-colored singleton was specifically suppressed while the target-colored and neutral-colored singletons were facilitated.
Salience and suppression
One might wonder whether a color singleton among three homogeneous grays in our paradigm would be salient enough to generate capture, as salience decreases as the number of homogeneous background items decreases (e.g., Leonard & Egeth, 2008; Meeter & Olivers, 2006; Theeuwes, 2004). Indeed, Wang and Theeuwes (2020) recently showed that larger displays (six or more items) render both target and distractor salient, resulting in attentional capture by the distractor even when participants engage in a feature-search mode. Those authors used a paradigm similar to that of Gaspelin et al. (2015) and compared singleton-suppression effects among set sizes of four, six, and ten. They found a singleton-suppression effect on probe trials when four items were presented, but the effect was reversed to enhanced processing of the singleton distractor when six or ten items were presented. Based on these findings, the authors concluded that signal suppression is limited to four-item displays. However, note that some other studies have found evidence consistent with inhibition of salient items (i.e., no attentional capture) in displays of six or more items (Feldmann-Wüstefeld et al., 2020; Gaspar & McDonald, 2014; Gaspelin et al., 2015, 2017; Sawaki & Luck, 2010).
In the current experiment, on search trials we successfully replicated the singleton presence benefit and, more importantly, on probe trials we found significant singleton-facilitation effects when a neutral-colored singleton (37 ms) or a target-colored singleton (65 ms) was presented. Even though we used four-item displays, the magnitudes of attentional capture on our neutral-colored singleton and target-colored singleton trials were comparable to or even larger than those in Wang and Theeuwes’ (2020) set size 6 (17 ms) or set size 10 (43 ms) experiments. The fact that we found such a substantial capture effect should not be surprising. On our probe trials all items were the same shape (ovals), and the singleton was a color among other items that were all the same shade of gray, which maximizes pop-out (Duncan & Humphrey, 1989; Nothdurft, 1993). We showed that this otherwise strongly salient item was suppressed when it was in the distractor color, which clearly demonstrates that a salience signal can be suppressed. Note that as the two paradigms use different procedures for calculating capture (see Footnote 3), our comparison of capture effects in the present experiment and in the Wang and Theeuwes (2020) experiment should be considered suggestive rather than definitive.
Be that as it may, the Wang and Theeuwes (2020) article is an important reminder that salience can vary with display size and that the relative salience of items in a display may be a critical factor in attentional selection. However, there may be factors in addition to display size that influence salience. For example, we have wondered whether colored outline shapes of the kind used by Wang and Theeuwes (2020) may lack the “oomph” of the large filled shapes used in Gaspelin et al. (2015) as well as in the present study (cf. Chang, Cunningham, & Egeth, 2019). Recently, Feldmann-Wüstefeld et al. (2020) also used outline shapes to examine whether irrelevant-but-salient features can be actively suppressed. In their study, the display consisted of one diamond-shaped (target) and five circular (distractor) outlines, each of which contained a digit. Participants were asked to report the digit inside the target. On singleton-absent trials, all items were gray outlines. On singleton-present trials, one of the distractors was a colored outline (i.e., a color singleton). Responses were slower on singleton-present trials than on singleton-absent trials, suggesting that the presence of salient items induced attentional costs. This finding is consistent with the finding from Wang and Theeuwes (2020). However, the singleton identity (i.e., the digit inside the color singleton) was less frequently reported than that of a nonsalient distractor, suggesting that a singleton distractor was suppressed. Furthermore, they found a decreased Pd component in the EEG signal preceding an erroneous report of the singleton identity. Since the Pd component is typically assumed to be a neural measure of suppression, this finding was taken as the evidence for active suppression of a singleton distractor. The authors argued that the necessary suppression that is applied to avoid attentional capture may be time demanding, thus resulting in longer reaction times on singleton-present trials. The absence of a distractor-N2pc that would be indicative of attentional capture supported an account in terms of nonspatial filtering costs (Folk & Remington, 1998). Without the “oomph” of large filled shapes, it may be possible that the presence of the color singleton slows down attentional allocation to the target without actually attracting attention; however, in a second step, suppression is applied to the color singleton.
Can incentive-salience be suppressed?
Suppression has proven itself to be a powerful competitor in the battle for attentional selection. In particular, we have shown that suppression can overcome capture by a salient color singleton. However, non-visual factors may also be relevant. Some research has explored this in the context of value-modulated attentional capture (VMAC, also referred to as VDAC, or value-driven attentional capture).
VMAC is typically studied in a two-phase experiment. For example, in Anderson, Laurent, and Yantis (2011), participants searched for a target circle that was either red or green in a display of chromatically heterogeneous nontarget circles. There was just one target item in each display and participants had to indicate the orientation of a line inscribed in the target item. Some participants received a large monetary reward when the target was red and a small reward when the target was green, while other participants had the reverse target-reward association. In the second phase of the experiment, no reward was provided. The displays were once again chromatically heterogeneous, but color was irrelevant; participants searched for a diamond in a display of multiple circles and again reported the orientation of an inscribed line. The crucial result was that when one of the non-target circles in the display was in the color of the high-value target from the first phase, reaction time to find the diamond was significantly slower than when the low-value target color was present. That is, the tendency to capture attention was modulated by reward value.
Summarizing some of the early work on this topic, Pearson, Watson, Cheng, and Le Pelley (2020) wrote, “…these findings suggest that the attentional priority of a reward-related stimulus is increased (at least in part) through the augmentation of its stimulus-driven perceptual activity, akin to increasing its incentive salience (Berridge & Robinson, 1998).” This leads one rather naturally to wonder whether this enhanced salience can be suppressed. Pearson et al. (2020) modeled their study after previous work by Gaspelin et al. (2015, 2017), who showed that physically salient stimuli could be suppressed when participants were induced to adopt feature-search mode, but not when they were induced to adopt singleton search mode (Bacon & Egeth, 1994). Pearson et al. (2020) showed that physically salient color stimuli associated with low reward could be suppressed, but that equally salient stimuli associated with high reward could not be suppressed, even when participants adopted feature-search mode. Note that in the Pearson et al. experiment the effects of distractor suppression and target enhancement were both operative, just as in Gaspelin et al.’s (2015, 2017) study. Thus, if the effect of target enhancement were eliminated, suppression alone would, a fortiori, not be sufficient to overcome the distracting effect of salience induced by high reward.
Grégoire, Britton, and Anderson (2020, Exp. 1) also attempted to determine if reward salience (of both positive and negative valence) could be suppressed. In the first phase participants saccaded to a shape oddball. One singleton color was associated with a monetary reward and another was associated with a small electrical shock; a third singleton color was associated with neither reward nor punishment. Participants were told that they were more likely to be rewarded if their response was fast and more likely to be shocked if their response was slow. Note that if participants can avoid wasting time making an eye movement to a distracting singleton they are more likely to be rewarded. In the test phase participants received neither reward nor punishment. The results indicated that participants could suppress highly valent singletons. For example, the proportion of first saccades to the distractor was significantly greater in the neutral condition than in either the reward or shock conditions, with no difference between the rewarded and shock condition. In a second experiment they eliminated electrical shock, meaning that there were, in effect, two neutral colors. Interestingly, they once again found evidence that participants could avoid looking at a previously highly rewarded stimulus when it was to their advantage to do so (see also Feldmann-Wüstefeld, Brandhofer, & Schubö, 2016). Grégoire et al. (2020) argued that with sufficient motivation value- and threat-modulated attentional capture can be suppressed. It is not yet clear what specific condition may have engendered higher motivation in the Gregoire et al. study than in the Pearson et al. study. In any case, it would be interesting to determine if suppression alone (without target enhancement) is sufficient to yield this result with high motivation.
Conclusion
The current study provides evidence that the strongest stimulus in a scene (here, a salient color singleton among homogeneous gray items) does not always automatically capture attention; it can be suppressed. This finding is consistent with the signal-suppression hypothesis, providing strong evidence that a salience signal generated by a salient stimulus can be actively suppressed even without target enhancement. Even though the signal-suppression hypothesis may not fully resolve the debate between stimulus-driven and goal-driven accounts of attentional selection (Wang & Theeuwes, 2020), suppression is still a potent mechanism that can override capture by salient-but-irrelevant singletons.
Notes
With a heterogeneous shape display, this task may have induced feature-search mode (e.g., Bacon & Egeth, 1994). In feature-search mode, the absence of a singleton capture effect is not surprising. However, the actual reversal of the capture effect is a striking finding.
We use the term “neutral” to refer to a singleton color that has not been used on search trials. Of course, in ordinary usage gray could also be considered neutral, but it doesn’t ever serve as a singleton. Please note the terminology is slightly different from that in Chang and Egeth (2019).
In classical paradigms, singleton capture effects are obtained when there is a singleton distractor slowing target search (e.g., Theeuwes, 1992). However, on probe trials in our paradigm, a singleton item on each display was either a target or a distractor item. Therefore, we refer to the obtained singleton related effects as either a singleton-facilitation effect or a singleton-inhibition effect depending on the pattern of the data.
References
Anderson, B. A., Laurent, P. A., & Yantis, S. (2011). Value-driven attentional capture. Proceedings of the National Academy of Sciences of the United States of America, 108(25), 10367–10371. https://doi.org/10.1073/pnas.1104047108
Awh, E., Belopolsky, A. V., & Theeuwes, J. (2012). Top-down versus bottom-up attentional control: A failed theoretical dichotomy. Trends in Cognitive Sciences, 16(8), 437–443. https://doi.org/10.1016/j.tics.2012.06.010
Bacon, W. F., & Egeth, H. E. (1994). Overriding stimulus-driven attentional capture. Perception & Psychophysics, 55(5), 485–496. https://doi.org/10.3758/BF03205306
Berridge, K. C., & Robinson, T. E. (1998). What is the role of dopamine in reward: Hedonic impact, reward learning, or incentive salience? Brain Research Reviews, 28(3), 309–369. https://doi.org/10.1016/S0165-0173(98)00019-8
Brainard, D. H. (1997).The Psychophysics Toolbox. Spatial Vision, 10, 433–436. https://doi.org/10.1163/156856897X00357
Chang, S., Cunningham, C. A., & Egeth, H. E. (2019). The power of negative thinking: Paradoxical but effective ignoring of salient-but-irrelevant stimuli with a spatial cue. Visual Cognition, 27(3–4), 199–213. https://doi.org/10.1080/13506285.2018.1541950
Chang, S., & Egeth, H. E. (2019). Enhancement and Suppression Flexibly Guide Attention. Psychological Science, 30(12), 1724–1732. https://doi.org/10.1177/0956797619878813
Cousineau, D. (2005). Confidence intervals in within-subject designs: A simpler solution to Loftus and Masson’s method. Tutorials in Quantitative Methods for Psychology, 1(1), 42–45. https://doi.org/10.20982/tqmp.01.1.p042
Duncan, J., & Humphrey, G. W. (1989). Visual serach and stimulus similarity. Psychological Review, 96(3), 433–458.
Feldmann-Wüstefeld, T., Brandhofer, R., & Schubö, A. (2016). Rewarded visual items capture attention only in heterogeneous contexts. Psychophysiology, 53(7), 1063–1073. https://doi.org/10.1111/psyp.12641
Feldmann-Wüstefeld, T., Busch, N. A., & Schubö, A. (2020). Failed suppression of salient stimuli precedes behavioral errors. Journal of Cognitive Neuroscience, 32(2), 367–377. https://doi.org/10.1162/jocn_a_01502
Folk, C. L., & Remington, R. (1998). Selectivity in distraction by irrelevant featural singletons: Evidence for two forms of attentional capture. Journal of Experimental Psychology: Human Perception and Performance, 24(3), 847–858. https://doi.org/10.1037//0096-1523.24.3.847
Folk, C. L., Remington, R. W., & Johnston, J. T. (1992). Involuntary Covert Orienting Is Contingent on Attentional Control Settings.Pdf.
Gaspar, J. M., & McDonald, J. J. (2014). Suppression of salient objects prevents distraction in visual search. Journal of Neuroscience, 34(16), 5658–5666. https://doi.org/10.1523/JNEUROSCI.4161-13.2014
Gaspelin, N., Gaspar, J. M., & Luck, S. J. (2019). Oculomotor inhibition of salient distractors: Voluntary inhibition cannot override selection history. Visual Cognition, 27(3–4), 227–246. https://doi.org/10.1080/13506285.2019.1600090
Gaspelin, N., Leonard, C. J., & Luck, S. J. (2015). Direct evidence for active suppression of salient-but-irrelevant sensory inputs. Psychological Science, 26(11), 1740–1750. https://doi.org/10.1177/0956797615597913
Gaspelin, N., Leonard, C. J., & Luck, S. J. (2017). Suppression of overt attentional capture by salient-but-irrelevant color singletons. Attention, Perception, and Psychophysics, 79(1), 45–62. https://doi.org/10.3758/s13414-016-1209-1
Gaspelin, N., & Luck, S. J. (2018a). Combined Electrophysiological and Behavioral Evidence for the Suppression of Salient Distractors. Journal of Cognitive Neuroscience, 30(9), 1265–1280. https://doi.org/10.1162/jocn
Gaspelin, N., & Luck, S. J. (2018b). Distinguishing Among Potential Mechanisms of Singleton Suppression Distinguishing Among Potential Mechanisms of Singleton Suppression. Journal of Experimental Psychology: Human Perception and Performance, 44(4), 626–644. https://doi.org/10.1037/xhp0000484
Grégoire, L., Britton, M. K., & Anderson, B. A. (2020). Motivated Suppression of Value- and Threat-Modulated Attentional Capture. Emotion. https://doi.org/10.1037/emo0000777
Hickey, C., Di Lollo, V., & McDonald, J. J. (2009). Electrophysiological indices of target and distractor processing in visual search. Journal of Cognitive Neuroscience, 21(4), 760–775. https://doi.org/10.1162/jocn.2009.21039
Itti, L., & Koch, C. (2000). A saliency-based search mechanism for overt and covert shifts of visual attention. Vision Research, 40(10–12), 1489–1506. https://doi.org/10.1016/S0042-6989(99)00163-7
Leonard, C. J., & Egeth, H. E. (2008). Attentional guidance in singleton search: An examination of top-down, bottom-up, and intertrial factors. Visual Cognition, 16(8), 1078–1091. https://doi.org/10.1080/13506280701580698
Meeter, M., & Olivers, C. N. L. (2006). Intertrial priming stemming from ambiguity: A new account of priming in visual search. Visual Cognition, 13(2), 202–222. https://doi.org/10.1080/13506280500277488
Nothdurft, H. C. (1993). The role of features in preattentive vision: Comparison of orientation, motion and color cues. Vision Research, 33(14), 1937–1958. https://doi.org/10.1016/0042-6989(93)90020-W
Pearson, D., Watson, P., Cheng, P. X., & Le Pelley, M. E. (2020). Overt Attentional Capture by Reward-Related Stimuli Overcomes Inhibitory Suppression. Journal of Experimental Psychology: Human Perception and Performance, 46(5), 489–501. https://doi.org/10.1037/xhp0000728
Sawaki, R., & Luck, S. J. (2010). Capture versus suppression of attention by salient singletons: Electrophysiological evidence for an automatic attend-to-me signal. Attention, Perception, & Psychophysics, 71(1), 174–182. https://doi.org/10.3758/APP
Theeuwes, J. (1992). Perceptual selectivity for color and form. Perception & Psychophysics, 51(6), 599–606. https://doi.org/10.3758/BF03211656
Theeuwes, J. (2004). Top-down search strategies cannot override attentional capture. Psychonomic Bulletin and Review, 11(1), 65–70. https://doi.org/10.3758/BF03206462
Theeuwes, J. (2018). Visual Selection: Usually Fast and Automatic; Seldom Slow and Volitional. Journal of Cognition, 1(1), 1–15. https://doi.org/10.5334/joc.13
Theeuwes, J. (2019). Goal-driven, stimulus-driven, and history-driven selection. Current Opinion in Psychology, 29, 97–101. https://doi.org/10.1016/j.copsyc.2018.12.024
Van Selst, M., & Jolicoeur, P. (1994). A Solution to the Effect of Sample Size on Outlier Elimination. The Quarterly Journal of Experimental Psychology Section A, 47(3), 631–650. https://doi.org/10.1080/14640749408401131
Wang, B., & Theeuwes, J. (2020). Salience determines attentional orienting in visual selection. Journal of Experimental Psychology. Human Perception and Performance. 46(10), 1051–1057. https://doi.org/10.1037/xhp0000796
Wyble, B., Callahan-Flintoft, C., Chen, H., Marinov, T., Sarkar, A., & Bowman, H. (2020). Understanding Visual Attention With RAGNAROC: A Reflexive Attention Gradient Through Neural AttRactOr Competition. Psychological Review. https://doi.org/10.1037/rev0000245
Acknowledgement
We thank Corbin A. Cunningham for valuable discussion and helpful comments.
Funding
This research was supported in part by a grant from the Kwanjeong Lee Chong Hwan Educational Foundation (KEF-2016) to S. Chang.
Author information
Authors and Affiliations
Corresponding author
Additional information
Open Practices Statement
The data for the experiment reported here are available at https://osf.io/f7np4 the experiment was not preregistered.
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Chang, S., Egeth, H.E. Can salient stimuli really be suppressed?. Atten Percept Psychophys 83, 260–269 (2021). https://doi.org/10.3758/s13414-020-02207-8
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13414-020-02207-8