
Abstract
Integrating information across the visual field into an ensemble (e.g., seeing the forest from the trees) is an effective strategy to efficiently process the visual world, and one that is often impaired in autism spectrum disorder. Individual differences in sensory processing predict ensemble encoding, providing a potential mechanism for differing perceptual strategies across individuals, and possibly across diagnostic groups exhibiting atypical sensory processing. Here, we explore whether ensemble encoding is associated with traits associated with autism spectrum disorder (ASD). Participants (N=68) were presented with an ensemble display consisting of circles of varying sizes and colors, and were asked to remember the size of the red and blue circles, while ignoring the green circles. Participants were then cued to a target location after a brief delay, and instructed to report the remembered size of the circle they had previously viewed in that location, as ensemble information commonly biases memory for individual objects toward the probed mean of a set of similar objects. The Autism-spectrum Quotient (AQ) was completed to measure each individual’s level of autistic traits. We found that an individual’s level of ensemble perception, measured as their bias toward the probed mean, was negatively associated with a higher level of ASD traits. These results suggest that individuals with higher levels of ASD traits are less likely to integrate perceptual information. These findings may shed light on different perceptual processing within the autism spectrum, and provide insight into the relationship between individual differences and ensemble encoding.
Similar content being viewed by others

Introduction
Our typical visual environment is overflowing with complexity, yet we still efficiently extract the gist of it, presumably by grouping redundant information. Specifically, it has been hypothesized that observers represent a set of similar items as an averaged group or ensemble, enabling the condensing of multiple individual elements into a single, higher level of description (Alvarez, 2011). This process would entail perceptually extracting a course-grained representation of the visual world (e.g., forest), rather than the fine-grained details of each object (e.g., individual trees). Prior research has demonstrated that ensemble, or summary, statistics implicitly bias the representation of individual items held in visual working memory, such that the remembered size of an individual item was biased toward the probed mean size of items sharing the same color (Brady & Alvarez, 2011). Given the capacity limits of visual working memory, this processing strategy may be critical for capturing the richness of our perceptual experience (Cohen et al., 2016). Recently, the integration of ensemble statistics with items held in visual working memory has been shown to correlate with individual differences in sensory processing patterns (Lowe et al., 2016b), which capture variability in behavioral strategies and neurological responses to general sensory experience (Dunn, 1997). Specifically, individuals with lower sensory thresholds were less likely to integrate mean size information across a set of similar items, suggesting a possible mechanism through which disordered perception may occur.
A growing body of evidence has shown that sensory processing abnormalities are inherently pervasive in individuals diagnosed with autism spectrum disorder (ASD) (Kern et al., 2006; for reviews, see Baum et al., 2015 and Simmons et al., 2009), suggesting that autistic traits may influence the processing of ensemble information from the visual environment. Many theories have emerged that may explain different perceptual processing in individuals with ASD, such as the weak central coherence cognitive theory (Frith, 1989), the temporal binding hypothesis (Brock et al., 2002), the theory of imbalanced excitation and inhibition (Rubenstein & Merzenich, 2003), and a Bayesian observer theory (Pellicano & Burr, 2012). One of the earlier frameworks, the weak central coherence cognitive theory, called attention to the tendency for individuals with ASD to show a processing bias for featural and local information relative to the ability to extract overall gist information. Indeed, individuals within the autism spectrum demonstrate superior performance on tasks requiring detail-focused processing compared with typically-developed (TD) individuals (Happé & Frith, 2006). For instance, autistic individuals may fail to integrate multiple pieces of visual information across a unified context, instead making more accurate judgments of individual items than TD individuals (Happé, 1996). Since the representation of ensemble statistics is dependent on the ability to integrate visual information within the context of similar items, the perceptual differences associated with ASD could compromise the ability to represent ensemble information accurately.
The Bayesian observer framework, which builds on the tenets of weak central coherence, further predicts that autistic individuals may be less able to utilize summary statistics from scenes due to either a reduced ability to integrate prior experience with current perception or a reduced ability to learn the statistics of the environment (Pellicano & Burr, 2012; Powell et al., 2016; Lawson et al., 2017). A number of studies have shown that the ability to utilize ensemble representations is reduced in ASD individuals relative to TD controls. For instance, global perceptual averaging of ensemble information, such as color, may be compromised in individuals with ASD (Maul et al. 2016), and ASD individuals show a reduced ability to average a set of faces (Rhodes et al., 2014). Likewise, individuals with ASD relative to their typically developed peers exhibit a reduced ability to estimate the mean size of a set of 16 circles via a two-alternative forced-choice task (it should be noted that this comparison was not directly reported, but calculations based on reported data show an effect size of d = 0.61; Corbett, Venuti, & Melcher, 2016). A second study exploring mean size estimation in ASD showed mixed results. Participants were presented with an array of ten circles followed by a probe and asked if the probe was smaller or larger than the mean. Disrupted ensemble processing in ASD was observed when the distributions of sizes were highly heterogeneous (ten different dot sizes), but not when more homogeneous (two different dot sizes) (Van der Hallen et al., 2017). With that said, individuals with ASD have demonstrated similar ensemble processing of arrays of emotional faces (Karaminis et al., 2017). Thus, while the majority of studies show some disruption in ensemble processing in ASD, there is some variability in findings, making it critical to investigate further how ensemble processing interacts with the autism spectrum.
As a spectrum disorder, traits associated with ASD are seen in the general population, and the study of autistic traits in non-clinical samples affords a practical way to explore aspects of the autism spectrum, and is an approach that has successfully generalized experimental findings in TD to similar findings in ASD populations (Bölte et al., 2011; Chandler et al., 2007; Constantino et al., 2003; Stevenson et al., 2018a). The present study examines the relationship between autistic traits and the representation of ensemble statistics in visual working memory in a TD population by combining an ensemble processing task (Brady & Alvarez, 2011), which has been previously used to measure individual differences in perception (Lowe et al., 2016b), with the self-assessed Autism-spectrum Quotient (AQ) questionnaire (Baron-Cohen et al., 2001). In this ensemble paradigm, participants are presented with an array of circles of varying colors, and asked to remember the size of particular colors of circles while ignoring others. When probed to recall the size of a single circle, participants often show a bias toward the mean circle size of the probed color, reflecting ensemble processing of that color set of circles (Fig. 1). Examining the encoding of ensemble bias for size estimates allows us to explore automatic encoding of simple ensemble statistics, and whether individual differences in autism traits are associated with perceptual biases. We predicted that individuals with higher levels of ASD traits would be less likely to integrate mean ensemble information with individual items, manifesting as a decreased likelihood that their estimates for individual items will be biased toward the probed mean of an ensemble display.

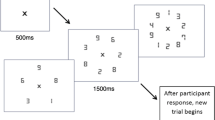
(a) Stimuli and trial order used in the ensemble statistics task. Participants were asked to remember the size of the red and blue circles, and ignore the green circles. After a short delay, they were asked to recall and select the size of a previously seen circle, which was cued by the presentation of a black probe circle. (b) Bias was determined from a pair of matched displays. Two matched displays had the same items, but the tested item’s colors (highlighted here with dashed lines) were swapped between the displays. Bias was calculated by dividing the reported size for probed items presented in the color of the larger circles by the reported size for probed items presented in the color of the smaller circles (see Brady & Alvarez, 2011). Note that the size of the circles is not to scale to illustrate how bias is determined
Method
Participants
Sixty-eight undergraduate psychology students in the University of Toronto community (18 male) ranging in age from 18 to 54 years (M = 21.3) participated and were compensated for participation with course credit. All participants had self-reported normal or corrected-to-normal visual acuity, no history of color-blindness or abnormal neurologic conditions, and gave informed consent in accordance with the University of Toronto Ethics Review Board.
Display and apparatus
The ensemble statistics task was presented electronically using Matlab (MathWorks, Natick, MA, USA), on a ViewSonic 21-in. CRT monitor (1,280 x 1,024; 85-Hz refresh rate) at a viewing distance of 57 cm (subtending 38.70° x 29.49° of visual angle) held constant by using a desk-mounted chinrest. The AQ was completed by pen and paper.
Ensemble statistics task
To examine the bias of ensemble statistics for a set of items sharing common features, we conducted a direct replication of the first experiment in Brady and Alvarez (2011).
Participants were presented with a total of 30 displays each consisting of nine circles of varying size (three red, three blue, and three green), and were asked to remember the size of the red and blue circles, while ignoring the green circles. By asking participants to ignore a subset of circles based on color, it was ensured that participants attended to the color of the items, as previous research has shown that when color was not task-relevant, ensemble biases are reduced or absent. That is, when only two sets of colored circles are presented in the array, participants can simply remember the size of all items, and thus fail to show a bias toward the probed mean as a function of color (Brady & Alvarez, 2011; Huang et al., 2007). It should also be noted that previous research has demonstrated that the inclusion of a third, unattended color set does not influence estimations of the attended color sets as long as participants are instructed as to which colors they must attend to in an a priori manner (Halberda et al., 2006). Each display appeared on screen for 1.5 s and was then followed by a 1-s blank period, after which participants were cued to a target location by a randomly-sized circle appearing in black at the location of a previously presented blue or red circle. Participants were then instructed to use the computer mouse to resize the black circle so that it matched the remembered size of the circle they had previously viewed in that location (see Fig. 1a).
The nine circles appeared on a gray background that measured 18° x 12° of visual angle, and each circle was randomly positioned in an unseen 6 x 4 grid, with a jitter of ± 0.3° of visual angle added to the circles’ locations to prevent collinearities. The mean diameter for the circles of each color (red, blue, and green) were randomly chosen from a uniform distribution ranging from 0.45° to 2.85° of visual angle. Each individual circle’s size was then chosen by sampling randomly from a distribution with that color’s chosen mean and an SD equal to one-eighths of this mean.
To directly test bias in reported size, 15 displays were generated, and then another 15 matched displays were created by switching the color of the to-be-tested item with the other non-distractor color (red and blue). Thus, two matched displays had the same items, but the tested item colors were swapped between the displays. The 30 displays were then randomly interleaved, with the constraint that paired displays could not appear one after the other.
Autism-spectrum quotient (AQ)
As a spectrum disorder, specific traits associated with ASD are not only found in individuals with a clinical diagnosis, but are also found in the general population. Such traits are commonly measured by using the Autism Quotient Questionnaire (AQ; Baron-Cohen et al. 2001), a 50-item, self-report measure, which all participants completed. The AQ has high internal consistency (Cronbach’s α = .97) and test-retest reliability (r = .85). The AQ has been known to reliably discriminate between individuals with and without ASD (sensitivity = 95%, specificity = 95%) at a cut-off score of 34. All participants were below this cut-off and reflected distributions previously collected with typically-developed samples (Baron-Cohen et al. 2001; Stevenson et al., 2017) (mean = 21.14, SD = 8.07, range = 9 to 31).
Results
Ensemble bias
To test ensemble bias, we examined whether participants’ size estimates were biased toward or away from the mean size representation of circles within the same color set. Each matched pair (consisting of two displays that had the same items, but the tested item’s colors were swapped between the displays) was divided on the basis of which of the pair contained a tested item the same color as the circles that were smaller on average and which contained a tested item the same color as the circles that were larger on average:
A ratio of 1.0 would indicate participants’ size estimates were not biased. However, if participants’ size estimates were biased toward the probed mean size of the circles in the same color as the tested item, this ratio would be greater than 1.0 (see Fig. 1B), with a maximum bias calculated as the value if each participant reported the ensemble mean itself of 1.481. This latter outcome was predicted based on previous evidence (Brady & Alvarez, 2011). On average, the reported size of the tested circle was 1.03 times greater on trials with the larger same-colored circles than on trials with the smaller same-colored circles (Fig. 2a), and this ratio was significantly greater than 1.0 (t(67) = 2.00, p = 0.05, d = 0.49). As is commonly found, variation in the directionality and extent of ensemble bias across participants was observed (see Fig. 2b), and we explore the source of this variation below.

(a) Mean bias averaged across participants. (b) The distribution and direction of mean size bias for all participants. For both graphs, a bias greater than 1.00 represents a bias toward the probed mean, and a bias less than 1.00 represents a bias away from the mean. Error bars represent the standard error of the mean. *p = 0.05
Ensemble bias and ASD traits
We first examined whether demographic variables (sex; age) correlated with ensemble bias. This investigation revealed no significant correlations (Pearson; two-tailed) between ensemble bias and either sex (r(68) = -0.034, confidence interval (CI) = [-0.270, 0.206], p = 0.782) or age (r(68) = -0.049, CI = [-0.284, 0.191], p = 0.691). To investigate our main hypothesis, we examined the correlation between ensemble bias and level of ASD traits as measured by the AQ (Fig. 3). We found a significant correlation between these two scores (r(68) = -0.269, CI = [-0.476, -0.033], p = 0.027. These results reveal a significant inverse relationship between ensemble bias and ASD traits; higher levels of ASD traits were associated with a decreased bias toward the probed mean ensemble representation. In contrast, no relationship was observed between ensemble bias and demographic factors.

Scatter plot (with best-fitting regression line) showing the correlation between ensemble bias (x-axis) and the autism-spectrum quotient (y-axis). An ensemble bias greater than 1.00 represents a bias toward the probed mean, and a bias less than 1.00 represents a bias away from the mean. CI confidence interval
We also conducted a multiple linear regression analysis to confirm a significant relationship between these two factors when demographic variables were accounted for, to investigate whether AQ scores could still predict ensemble bias. Indeed, this analysis revealed that scores on the AQ significantly predicted ensemble bias (t(67) = 2.34, p = 0.022, d = 0.57) with both sex (t(67) = 0.43, p = 0.668, d =0.11) and age (t(67) = 0.70, p = 0.484, d =0.17) accounted for.
Finally, a number of analyses were conducted to ensure that extraneous variability did not account for these reported effects. First, with randomly selected stimulus sizes on each trial, the variability of size within the target set on each trial also randomly varied. To ensure that this variability did not affect our results, we calculated the variance within each target ensemble and correlated this variance with ensemble bias, finding no relationship between the two (r = -0.007, p = 0.82). Second, the task-irrelevant green circle sizes also varied randomly. A mixed linear model was conducted to measure the contribution that the mean size of each ensemble, in addition to the target items size, contributed to participants’ responses, while controlling for the demographic variables of age, gender, and IQ. As expected, the target item’s size was the most significant predictor of participants’ response (t(1009) = 10.07, p < 0.001), followed by the target ensembles mean size (t(1000) = 2.26, p = 0.02). Neither the mean size of the distractor ensemble (t(1005) = 0.32, p = 0.75) nor the mean size of the task-irrelevant ensemble (t(1010) = -1.23, p = 0.22) significantly contributed to participant responses. Likewise, no demographic variable, including age (t(67) = 0.47, p = 0.64), gender (t(67) = -0.45, p = 0.66), or IQ (t(67) = 0.36, p = 0.72), contributed significantly to the model.
Discussion
Sensory and perceptual issues associated with autism have garnered a great deal of research attention, with a primary hypothesis that individuals with ASD often exhibit difficulties integrating individual pieces of sensory information into a single, unified perception. Our current findings test this hypothesis using a particular instantiation of this processes, namely ensemble perception, and show that individuals with higher levels of autistic traits show decreased ensemble perception. This relationship holds even when controlling for demographic factors (i.e., age, sex). Specifically, individuals with levels of ASD traits were less likely to represent individual items of a certain color as biased toward the probed mean of an ensemble representation with items containing that color, and thus were less likely to integrate similar perceptual information from the environment. These results are consistent with previous findings linking sensory processing patterns with measures of ensemble bias (Lowe et al., 2016b), yet extend this work by providing a more direct investigation of the relationship between ensemble processing and autistic traits.
The visual world is crowded with objects sharing similar attributes (e.g., faces in a crowd; trees in a forest), and ensemble encoding can be used to extract a tremendous amount of redundant information to obtain an almost immediate sense of the visual environment (Cohen et al., 2016). In fact, neural regions involved in the processing of ensemble information overlap with scene-selective areas (Cant & Xu, 2012, 2015), and these areas may represent multiple properties of the visual environment that influence our perceptual experience of the world (Lowe et al., 2016a). The present findings may therefore shed light on the cognitive mechanisms underlying the different, and often detail-focused, experience of the visual world observed in individuals with ASD, and also build on existing evidence demonstrating how individual differences influence the processing of ensemble features in the environment (Yang et al., 2013; Haberman, Brady, & Alvarez, 2015; Lowe et al., 2016b).
Previous research has suggested a decreased ability of individuals with ASD to integrate multiple pieces of information into a coherent Gestalt more broadly (Frith, 1989), particularly pertaining to the integration of sensory information (Baum et al., 2015; Stevenson et al. 2014b; Wallace and Stevenson, 2014), as such disturbances in sensory integration have been linked to symptomology in ASD (Stevenson et al., 2014a, 2018b; Woynaroski et al., 2013). This impaired ability to integrate global information across a unified context has been pivotal to theoretical accounts of ASD, particularly the weak central coherence theory (Frith, 1989), yet there has been much debate about this claim. In fact, there is growing support that, while individuals with ASD may show a preference for local information, they do not show an impairment in the ability to process global information (Koldewyn et al., 2013; Van der Hallen, 2015). In a similar vein, the integration of current sensory inputs and prior experiences has been demonstrated to be reduced in ASD (Powell et al., 2016; Karaminis et al., 2016; Lawson et al., 2017; Noel et al., 2017, 2018; Sevgi, Diaconescu, Tittgeeyer, & Schilback, 2016, 2012; Turi et al., 2015; Stevenson et al., 2017). Specifically, it has been proposed that these findings reflect difficulties in the ability to extract summary statistics, difficulties in the ability to integrate sensory representations, or both (Maul, Stanworth, Pellicano, & Franklin, 2016). Our data here, showing that individuals with high levels of ASD traits exhibit a decreased influence of ensemble statistics, which are by nature reliant upon the integration of information across individual item presentations, support these hypotheses.
Individuals with ASD are able to represent some level of ensemble information, but this representation may be compromised when compared to TD individuals (Corbett, Venuti, & Melcher, 2016; Maul, Stanworth, Pellicano, & Franklin, 2016; Rhodes, Neumann, Ewing, & Palermo, 2014), though experimental variables such as ensemble heterogeneity may influence this effect (Van der Hallen et al., 2017). The results of the present study expand these latter findings to suggest that these findings are not constrained to individuals with a clinical diagnosis, but can also be observed in the general population relative to their individual level of autistic traits. The precise mechanisms underlying this relationship are presently unclear. One suggestion previously put forth suggests a reduced ability to integrate prior experience with current perception may affect the extraction of ensemble statistics in ASD individuals (Pellicano & Burr, 2012). Individual differences in sensory processing patterns predict the bias of ensemble statistics held in visual working memory (Lowe et al., 2016b). Since ASD traits as measured by the AQ have been shown to correlate significantly with sensory scales (Black et al., 2017; Holder, Wilson, Mendez, & Murphy, 2014; Schulz & Stevenson, in press), the reduced ensemble bias in individuals with high AQ scores seen here may be related to general sensory experience. Other mechanisms, including but not limited to global versus local deployment of attention, the ability to supress distracting information, and the ability to form perceptual gestalts for similar items, may also influence this relationship. These possibilities should be explored in greater detail in future studies. Moreover, since the present study investigated TD individuals, future research should also explore whether individuals clinically diagnosed with ASD exhibit similar patterns of ensemble processing, through tasks assessing not only the overall ability to represent ensemble information, but also the sensitivity to ensemble information across multiple sensory domains, both explicit and implicit.
It should also be noted that our results here pertain directly to ensemble processing of the size of objects in an array, and these results may not generalize to ensemble processing of all types of stimulus features. Though these results are similar to previous findings of decreased ensemble processing of size, color, and facial identity (Corbett et al. 2016; Maul et al., 2016; Rhodes et al., 2014; Van der Hallen et al., 2017), this decrease was not observed with ensemble processing of emotional expression (Karaminis et al., 2017) or with more homogeneous arrays (Van der Hallen et al., 2017). As such, future research incorporating multiple measures of ensemble processing across an array of stimulus features would be quite beneficial.
In summary, our findings reveal a relationship between autistic traits as measured by the AQ and the bias of ensemble statistics for perceptually similar information held in visual working memory. These findings may shed light on different perceptual processing within the autism spectrum, and provide insight into the relationship between individual differences and ensemble encoding.
References
Alvarez, G. A. (2011). Representing multiple objects as an ensemble enhances visual cognition. Trends in cognitive sciences, 15(3), 122-131.
Baron-Cohen, S., Wheelwright, S., Skinner, R., Martin, J., & Clubley, E. (2001). The autism-spectrum quotient (AQ): Evidence from asperger syndrome/high-functioning autism, males and females, scientists and mathematicians. Journal of autism and developmental disorders, 31(1), 5-17
Baum, S. H., Stevenson, R. A., & Wallace, M. T. (2015). Behavioral, perceptual, and neural alterations in sensory and multisensory function in autism spectrum disorder. Progress in neurobiology, 134, 140-160.
Black, K. R., Stevenson, R. A., Segers, M., Ncube, B. L., Sun, S. Z., Philipp-Muller, ... Ferber, S. (2017). Linking Anxiety and Insistence on Sameness in Children with Autism: The Role of Hypersensitivity. Journal of Autism and Developmental Disabilities. https://doi.org/10.1007/s10803-017-3161-x
Bölte, S., Westerwald, E., Holtmann, M., Freitag, C., & Poustka, F. (2011). Autistic traits and autism spectrum disorders: The clinical validity of two measures presuming a continuum of social communication skills. Journal of Autism and Developmental Disorders, 41(1), 66-72.
Brady, T. F., & Alvarez, G. A. (2011). Hierarchical encoding in visual working memory ensemble statistics bias memory for individual items. Psychological Science, 22(3), 384-392.
Brock, J., Brown, C. C., Boucher, J., & Rippon, G. (2002). The temporal binding deficit hypothesis of autism. Development and psychopathology 14(02), 209-224.
Cant, J. S., & Xu, Y. (2012). Object ensemble processing in human anterior-medial ventral visual cortex. The Journal of Neuroscience, 32(22), 7685-7700.
Cant, J. S., & Xu, Y. (2015). The impact of density and ratio on object-ensemble representation in human anterior-medial ventral visual cortex. Cerebral Cortex, 25(11), 4226-4239.
Cohen, M. A., Dennett, D. C., & Kanwisher, N. (2016). What is the bandwidth of perceptual experience? Trends in cognitive sciences, 20(5), 324-335.
Corbett, J. E., Venuti, P., & Melcher, D. (2016). Perceptual averaging in individuals with Autism Spectrum Disorder. Frontiers in Psychology, 7.
Chandler, S., Charman, T., Baird, G., Simonoff, E., Loucas, T., Meldrum, D., . . . Pickles, A. (2007). Validation of the social communication questionnaire in a population cohort of children with autism spectrum disorders. Journal of the American Academy of Child & Adolescent Psychiatry, 46(10), 1324-1332.
Constantino, J. N., Davis, S. A., Todd, R. D., Schindler, M. K., Gross, M. M., Brophy, S. L., ... Reich, W. (2003). Validation of a brief quantitative measure of autistic traits: comparison of the social responsiveness scale with the autism diagnostic interview-revised. Journal of Autism and Developmental Disorders, 33(4), 427-433.
Dunn, W. (1997). The impact of sensory processing abilities on the daily lives of young children and their families: A conceptual model. Infants & Young Children, 9(4), 23-35.
Frith, U. (1989). Autism: Explaining the enigma (Vol. 1989). Blackwell Scientific Publications: Oxford.
Haberman, J., Brady, T. F., & Alvarez, G. A. (2015). Individual differences in ensemble perception reveal multiple, independent levels of ensemble representation. Journal of Experimental Psychology: General, 144(2), 432.
Halberda, J., Sires, S. F., & Feigenson, L. (2006). Multiple spatially overlapping sets can be enumerated in parallel. Psychological science, 17(7), 572-576.
Happé, F. G. (1996). Studying weak central coherence at low levels: Children with autism do not succumb to visual illusions. A research note. Journal of Child Psychology and Psychiatry, 37(7), 873-877.
Happé, F., & Frith, U. (2006). The weak coherence account: Detail-focused cognitive style in autism spectrum disorders. Journal of autism and developmental disorders, 36(1), 5-25.
Horder, J., Wilson, C. E., Mendez, M. A., & Murphy, D. G. (2014). Autistic traits and abnormal sensory experiences in adults. Journal of Autism and Developmental Disorders, 44(6), 1461-1469.
Huang, L., Treisman, A., & Pashler, H. (2007). Characterizing the limits of human visual awareness. Science, 317(5839), 823-825.
Karaminis, T., Cicchini, G.M., Neil, L., Cappagli, G., Aagten-Murphy, D., Burr, D., & Pellicano, E. (2016). Central tendency effects in time interval reproduction in autism. Scientific Reports, 6, 28570.
Karaminis, T., Neil, L., Manning, C., Turi, M., Fiorentini, C., Burr, D., & Pellicano, E. (2017). Ensemble perception of emotions in children with autism is similar to typically developing children. Developmental Cognitive Neuroscience, 24, 51-62.
Kern, J. K., Trivedi, M. H., Garver, C. R., Grannemann, B. D., Andrews, A. A., Savla, J. S., ... & Schroeder, J. L. (2006). The pattern of sensory processing abnormalities in autism. Autism, 10(5), 480-494.
Koldewyn, K., Jiang, Y. V., Weigelt, S., & Kanwisher, N. (2013). Global/local processing in autism: Not a disability, but a disinclination. Journal of autism and developmental disorders, 43(10), 2329-2340.
Lawson, R. P., Mathys, C., & Rees, G. (2017). Adults with autism overestimate the volatility of the sensory environment. Nature neuroscience, 20(9), 1293.
Lowe, M. X., Gallivan, J. P., Ferber, S., & Cant, J. S. (2016a). Feature diagnosticity and task context shape activity in human scene-selective cortex. NeuroImage, 125, 681-692.
Lowe, M. X., Stevenson, R. A., Wilson, K. E., Ouslis, N. E., Barense, M. D., Cant, J. S., & Ferber, S. (2016b). Sensory processing patterns predict the integration of information held in visual working memory. Journal of Experimental Psychology: Human Perception and Performance, 42(2), 294.
Maule, J., Stanworth, K., Pellicano, E., & Franklin, A. (2016). Ensemble perception of color in autistic adults. Autism Research.
Noel, J. P., De Niear, M. A., Stevenson, R., Alais, D., & Wallace, M. T. (2017). Atypical rapid audio-visual temporal recalibration in autism spectrum disorders. Autism Research, 10(1), 121-129.
Noel, J. P., Stevenson, R. A., & Wallace, M. T. (2018). Atypical audiovisual temporal function in autism and schizophrenia: Similar phenotype, different cause. European Journal of Neuroscience, 47(10), 1230-1241.
Pellicano, E., & Burr, D. (2012). When the world becomes ‘too real’: A Bayesian explanation of autistic perception. Trends in cognitive sciences, 16(10), 504-510.
Powell, G., Meredith, Z., McMillin, R., & Freeman, T. C. (2016). Bayesian models of individual differences: combining autistic traits and sensory thresholds to predict motion perception. Psychological science, 27(12), 1562-1572.
Rhodes, G., Neumann, M.F., Ewing, L., & Palermo, R. (2014). Reduced set averaging of face identity in children and adolescents with autism. Quarterly Journal of Experimental Psychology, 68, 1394–1403.
Rubenstein, J. L. R., & Merzenich, M. M. (2003). Model of autism: Increased ratio of excitation/inhibition in key neural systems. Genes, Brain and Behavior, 2(5), 255-267.
Sevgi, M., Diaconescu, A.O., Tittgemeyer, M., & Schilbach, L. (2016). Social Bayes: Using Bayesian modeling to study autistic trait–related differences in social cognition. Biological Psychiatry, 80, 112–119.
Schultz, S. E., & Stevenson, R. A. (In Press). Sensory hypersensitivity predicts repetitive behaviors in autistic and typically-developed children. Autism.
Simmons, D. R., Robertson, A. E., McKay, L. S., Toal, E., McAleer, P., & Pollick, F. E. (2009). Vision in autism spectrum disorders. Vision research, 49(22), 2705-2739.
Stevenson, R. A., Siemann, J. K., Schneider, B. C., Eberly, H. E., Woynaroski, T. G., Camarata, S. M., & Wallace, M. T. (2014a). Multisensory temporal integration in autism spectrum disorders. The Journal of Neuroscience, 34(3), 691-697.
Stevenson, R. A., Segers, M., Ferber, S., Barense, M. D., & Wallace, M. T. (2014b). The impact of multisensory integration deficits on speech perception in children with autism spectrum disorders. Frontiers in Psychology, 249.
Stevenson, R. A., Toulmin, J. K., Youm, A., Besney, R. M., Schulz, S. E., Barense, M. D., & Ferber, S. (2017). Increases in the autistic trait of attention to detail are associated with decreased multisensory temporal adaptation. Scientific reports, 7(1), 14354.
Stevenson, R. A, Segers, M., Ncube, B. L., Black, K. R., Bebko, J. M., Ferber, S., & Barense, M. D. (2018a). The cascading influence of low-level multisensory processing on speech perception in autism. Autism, 22(5), 609-624.
Stevenson, R. A., Sun, S. Z., Hazlett, N., Cant, J. S., Barense, M. D., & Ferber, S. (2018b). Seeing the forest and the trees: default local processing in individuals with high autistic traits does not come at the expense of global attention. Journal of autism and developmental disorders, 1-15.
Turi, M., Burr, D.C., Igliozzi, R., Aagten-Murphy, D., Muratori, F., & Pellicano, E. (2015). Children with autism spectrum disorder show reduced adaptation to number. Proceedings of the National Academy of Sciences of the United States of the America, 112, 7868–7872
Van der Hallen, R., Evers, K., Brewaeys, K., Van den Noortgate, W., & Wagemans, J. (2015). Global processing takes time: A meta-analysis on local–global visual processing in ASD. Psychological bulletin, 141(3), 549.
Van der Hallen, R., Lemmens, L., Steyaert, J., Noens, I., & Wagemans, J. (2017). Ensemble perception in autism spectrum disorder: Member-identification versus mean-discrimination. Autism Research.
Wallace, M. T., & Stevenson, R. A. (2014). The construct of the multisensory temporal binding window and its dysregulation in developmental disabilities. Neuropsychologia, 64, 105-123.
Woynaroski, T. G., Kwakye, L. D., Foss-Feig, J. H., Stevenson, R. A., Stone, W. L., & Wallace, M. T. (2013). Multisensory speech perception in children with autism spectrum disorders. Journal of autism and developmental disorders, 43(12), 2891-2902.
Yang, J. W., Yoon, K. L., Chong, S. C., & Oh, K. J. (2013). Accurate but pathological: Social anxiety and ensemble coding of emotion. Cognitive Therapy and Research, 37(3), 572-578.
Acknowledgements
R.A.S is supported by internal grants from the University of Western Ontario, a Social Sciences and Humanities Research Council of Canada (SSHRC) Insight grant (R5502A07), an NSERC Discovery Grant (RGPIN-2017-04656), an Ontario Early Researcher Award, and the John R. Evans Leaders Fund from the Canadian Foundation for Innovation (#37497). J.S.C. is supported by a Natural Sciences and Engineering Research Council (NSERC) Grant (216203-13) and Canadian Institutes of Health Research (CIHR) Grant (106436) to S.F., an NSERC grant to J.S.C. (435647-13). M.D.B. is supported by the Canada Research Chairs Program and a James S. McDonnell Scholar Award. The authors thank Timothy Brady and George Alvarez at Harvard University for providing their ensemble experiment and analysis procedure.
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
Ethical approval
All procedures performed in studies involving human participants were in accordance with the ethical standards of the institutional and/or national research committee and with the 1964 Helsinki declaration and its later amendments or comparable ethical standards.
Informed consent
Informed consent was obtained from all individual participants included in the study.
Conflict of interest
All authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Additional information
Significance
Ensemble encoding is a perceptual strategy to process the visual world efficiently by extracting group level information without needing to process each individual item in the ensemble. This study explored whether ensemble encoding is associated with traits associated with autism, a disorder associated with atypical sensory processing, particularly in the realm of integrating multiple pieces of sensory information into a unified whole. We found that an individual’s level of ensemble perception was negatively associated with a higher level of autistic traits. These results suggest that individuals with higher levels of autistic traits are less likely to integrate perceptual information.
Rights and permissions
About this article

Cite this article
Lowe, M.X., Stevenson, R.A., Barense, M.D. et al. Relating the perception of visual ensemble statistics to individual levels of autistic traits. Atten Percept Psychophys 80, 1667–1674 (2018). https://doi.org/10.3758/s13414-018-1580-1
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13414-018-1580-1