
Abstract
A puzzling interaction involving alertness and cognitive control is indicated by the finding of faster performance but larger congruency effects on alert trials (on which alerting cues are presented before the task stimuli) than on no-alert trials in selective attention tasks. In the present study, the author conducted four experiments to test hypotheses about the interaction. Manipulation of stimulus spacing revealed a difference in congruency effects between alert and no-alert trials for narrowly spaced stimuli but not for widely spaced stimuli, inconsistent with the hypothesis that increased alertness is associated with more diffuse attention. Manipulation of color grouping revealed similar differences in congruency effects between alert and no-alert trials for same-color and different-color groupings of targets and distractors, inconsistent with the general hypothesis that increased alertness is associated with more perceptual grouping. To explain the results, the author proposes that increased alertness is associated specifically with more spatial grouping of stimuli, possibly by modulating the threshold for parsing stimulus displays into distinct objects.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
The human attention system can be partitioned into components associated with alertness, orienting, and cognitive control (Fan, McCandliss, Sommer, Raz, & Posner, 2002; Posner & Boies, 1971; Posner & Petersen, 1990). Alertness reflects temporary or sustained changes in sensitivity to external stimulation or readiness to respond. Orienting involves shifting attention toward specific objects or locations in the environment. Cognitive control is associated with mechanisms for focusing attention and resolving conflicts in information processing. These components of attention have been studied in isolation but also in combination, and the present study addresses a puzzling interaction involving alertness and cognitive control reported in previous work.
The interaction has been demonstrated in variants of the Attention Network Test (ANT) developed by Fan et al. (2002), which combines flanker (B. A. Eriksen & Eriksen, 1974) and cuing (Posner, 1980) tasks for studying attention. On each trial of the ANT, subjects indicate whether a target arrow presented above or below fixation is pointing left or right by making a spatially corresponding response. The target arrow is flanked horizontally by neutral stimuli or by distractor arrows associated with the same response as the target (congruent) or with the opposite response (incongruent). Congruency effects—longer response times (RTs) for incongruent than for congruent stimuli—are routinely found and interpreted as evidence of failures to attend exclusively to targets. Cognitive control is assumed to be invoked to address the limitations of selective attention, but there is evidence that it is modulated by alertness: Congruency effects are often larger for alert trials, on which stimuli are preceded by alerting cues (e.g., warning signals presented briefly at the potential target locations), than for no-alert trials (e.g., Fan et al., 2002; Redick & Engle, 2006; Schneider, 2018; for overviews, see MacLeod et al., 2010; Nieuwenhuis & de Kleijn, 2013). Notably, RTs are generally shorter on alert than on no-alert trials, suggesting that the modulation of cognitive control occurs in the context of increased alertness that facilitates overall responding.
At least five hypotheses have been proposed to explain the finding of faster performance but greater distractor influence on alert than on no-alert trials. One hypothesis is that increased alertness inhibits cognitive control to facilitate processing of sensory input and produce fast responses (Callejas, Lupiáñez, Funes, & Tudela, 2005; Callejas, Lupiáñez, & Tudela, 2004). Another hypothesis is that alerting cues modulate response selection by activating established stimulus–response associations, exacerbating interference for incongruent stimuli associated with competing responses (Böckler, Alpay, & Stürmer, 2011; Fischer, Plessow, & Kiesel, 2010, 2012). A third possibility is the early onset hypothesis, which is that increased alertness shortens stimulus encoding time, allowing response selection to start before sufficient time has elapsed for cognitive control processes to focus attention on the target (Nieuwenhuis & de Kleijn, 2013; Schneider, 2018). A fourth idea is the global processing hypothesis, which is that increased alertness enhances global processing of stimuli, resulting in more attention to distractors and less localized processing of targets (Weinbach & Henik, 2011, 2012).
A fifth possibility, which was the initial motivation for the present study, is what I call the diffuse attention hypothesis (McConnell & Shore, 2011; Weinbach & Henik, 2012). It is related to the early onset hypothesis, which involves the assumption that attention is more diffuse when response selection starts on alert than on no-alert trials. It is also related to the global processing hypothesis, by which alerting cues elicit a global processing bias by triggering a more diffuse attentional focus. Visual selective attention is often likened to a spotlight or zoom lens of adjustable size that can be directed to specific regions of the visual field (e.g., C. W. Eriksen & St. James, 1986; Heitz & Engle, 2007; LaBerge, 1983; for a review, see Cave & Bichot, 1999). Alerting cues might trigger widening of the attentional spotlight, especially on double-cue trials—where the alerting cues are visual signals appearing above and below fixation at the potential target locations—that were originally used to measure alerting effects in the ANT (Fan et al., 2002). Diffuse attention following alerting cues might benefit subsequent stimulus detection (shortening overall RTs) but allow for more distractor processing (increasing congruency effects).
The limited available evidence concerning the diffuse attention hypothesis is mixed. Weinbach and Henik (2011) indirectly tested the hypothesis using a global/local task paradigm (Navon, 1977). In their experiment, subjects saw a hierarchically organized stimulus display consisting of a left-pointing or right-pointing large arrow composed of smaller arrows that pointed in the same direction or in the opposite direction as the large arrow (congruent and incongruent stimuli, respectively). An auditory alerting cue sometimes preceded the stimulus display. In global-task blocks, in which subjects responded to the direction of the large arrow, similar congruency effects were obtained on alert and no-alert trials. In local-task blocks, in which subjects responded to the direction of the smaller arrows, a larger congruency effect was obtained on alert than on no-alert trials. The greater influence of the irrelevant global stimulus on local-task performance under increased alertness is consistent with a wider attentional spotlight encompassing the large arrow, providing indirect support for the diffuse attention hypothesis (but see Weinbach & Henik, 2014, for evidence that stimulus salience matters in their paradigm).
While the present manuscript was being written, I obtained a preprint of a study by Seibold (2018) that involved direct testing of the diffuse attention hypothesis. In her experiments, subjects performed a primary task in which they processed the identity of a central letter in a horizontal stimulus array. An auditory alerting cue sometimes preceded the stimulus display. Some trials also involved a secondary task for which subjects responded to a probe occurring at a random spatial position of the stimulus array. Because the primary task involved focusing attention at the center of the stimulus array, probe RTs were predicted to be shortest at that position and progressively longer at more distant positions, yielding a V-shaped probe-RT function reflecting an attentional gradient (LaBerge, 1983). If increased alertness results in more diffuse attention, then the probe-RT function should be flatter on alert than on no-alert trials. Across five experiments, Seibold obtained V-shaped probe-RT functions that were similar for alert and no-alert trials, inconsistent with the diffuse attention hypothesis.
Given the limited evidence concerning whether increased alertness is associated with a widened attentional spotlight, there is a need for further investigation of the diffuse attention hypothesis. It seems reasonable to assume that if congruency effects are larger on alert than on no-alert trials because of a wider attentional focus that encompasses more distractors, then the interaction should be sensitive to the spatial distance between stimuli. In the flanker task (essentially the ANT without alerting cues), congruency effects decrease as targets and distractors are spaced farther apart (B. A. Eriksen & Eriksen, 1974; Hübner, Steinhauser, & Lehle, 2010; Kramer & Jacobson, 1991; Mattler, 2006; Miller, 1991). The main explanation of this finding is that distant distractors are less likely to fall within the attentional spotlight, or they are allocated less attentional capacity as they get farther from the center of the spotlight (reflecting an attentional distribution or gradient; Cohen, Servan-Schreiber, & McClelland, 1992; LaBerge & Brown, 1989; White, Ratcliff, & Starns, 2011), thereby reducing distractor influence.
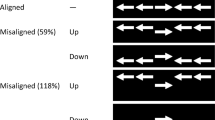
In Experiment 1, I manipulated stimulus spacing in a simplified variant of the ANT to test the diffuse attention hypothesis. Subjects classified the direction of a target arrow presented above or below fixation and flanked horizontally by incongruent or congruent distractor arrows. On no-alert trials, the arrow stimuli appeared after a variable fixation interval. On alert trials, alerting cues (squares at the potential target locations) were presented briefly in advance of the arrow stimuli during the fixation interval. The spacing of adjacent arrows was either narrow or wide on different trials (see Fig. 1).

I expected shorter overall RTs but larger congruency effects on alert than on no-alert trials (e.g., as in Fan et al., 2002; MacLeod et al., 2010; Schneider, 2018). I also expected congruency effects to decrease with wider spacing of stimuli (e.g., as in B. A. Eriksen & Eriksen, 1974; Miller, 1991). Critically, the diffuse attention hypothesis leads to the prediction of a three-way interaction between congruency, alerting, and spacing. As stimulus spacing increases, distractors are less likely to fall within the attentional spotlight; however, if alerting cues trigger widening of the spotlight, then distant distractors might enter the spotlight. The behavioral outcome would be a larger difference in congruency effects between alert and no-alert trials with wider spacing of stimuli. Framed another way, the decrease in the congruency effect with wider spacing would be attenuated on alert trials relative to no-alert trials. The occurrence of this three-way interaction would support the diffuse attention hypothesis.
Experiment 1
Method
Subjects
A total of 111 students from Purdue University participated for course credit. A simulation-based power analysis using alerting data from Schneider (2018) and spacing data from an unpublished experiment (without alerting) indicated that 104 subjects would provide 90% power to detect a three-way interaction for which the change in the congruency effect with wider spacing was halved on alert versus no-alert trials. The targeted sample size was exceeded because extra subjects were available during the final week of data collection. Data from one additional subject were excluded because of a failure to follow instructions (responses were made to distractors instead of targets) that resulted in a mean error rate (averaged over all conditions) greater than 20%. All subjects reported having normal or corrected-to-normal vision (there was no restriction about having normal color vision in this experiment).
Apparatus
The experiment was conducted using computers that displayed stimuli on monitors viewed from an uncontrolled distance of approximately 50 cm. Responses were registered from Chronos devices (Psychology Software Tools, Inc.). The leftmost and rightmost buttons on the response device were pressed with the left and right index fingers, respectively.
Task and stimuli
The task was to indicate whether a target arrow was pointing left or right by pressing a spatially corresponding response button. The target arrow was flanked horizontally by four identical distractor arrows (two on the left and two on the right) pointing either left or right, such that the distractors were associated with the same response as the target (congruent) or with the opposite response (incongruent). Each arrow measured 1.15° wide × 0.46° high, and the spacing of adjacent arrows (edge-to-edge distance) was either narrow (0.11°) or wide (1.26°; see Fig. 1). All stimuli were displayed in white on a black background.
Procedure
Subjects were tested individually after giving informed consent for a study protocol approved by the Purdue University Institutional Review Board. Instructions were presented onscreen and read aloud by the experimenter. Subjects were asked to classify quickly and accurately the direction of the middle arrow on each trial, ignoring the other arrows. During the instructions, subjects performed eight example no-alert trials (reflecting all combinations of congruency, spacing, and target direction) with accuracy feedback. Afterward, they performed nine blocks of 48 trials per block without accuracy feedback.
The procedures for no-alert and alert trials resembled those used for no-cue and double-cue trials, respectively, in the ANT (Fan et al., 2002). Each trial started with a central fixation cross (0.23° × 0.23°). On no-alert trials, after a randomly selected fixation interval of 1,000, 1,500, or 2,000 ms, arrow stimuli appeared 1.09° above or below fixation. The stimuli remained visible until the subject responded, then disappeared (leaving only the fixation cross) and the next trial started. On alert trials, the procedure was identical, except that 500 ms before the arrows appeared, alerting cues (filled squares measuring 0.46° × 0.46°) were presented for 100 ms above and below fixation at the potential target locations. Stimulus locations (above or below fixation) were randomly selected but occurred equally often across trials. Every possible combination of fixation interval, alerting, congruency, spacing, and target direction occurred equally often in random order in each block.
Results
The first block of trials was excluded as practice. Trials with RTs more than three standard deviations above the mean in each condition for a given subject were excluded (1.6% of trials). Error trials were excluded from the RT analysis. Mean RTs and error rates are reported as a function of congruency (incongruent or congruent), alerting (no alert or alert), and spacing (narrow or wide) in Table 1. The results of repeated-measures analyses of variance (ANOVAs) with those variables as factors are summarized in Table 2. The mean error rate was 2.3%, and there were no indications of speed–accuracy trade-offs; therefore, I focused on the RT data.
Performance was worse for incongruent stimuli (mean RT of 517 ms) than for congruent stimuli (457 ms), yielding a significant main effect of congruency. Subjects responded faster on alert trials (470 ms) than on no-alert trials (504 ms), yielding a significant main effect of alerting. Subjects were also faster when stimulus spacing was wide (471 ms) than when it was narrow (503 ms), yielding a significant main effect of spacing. Alerting effects (differences in RTs between no-alert and alert trials) increased from 32 ms to 36 ms with wider spacing, yielding a significant interaction between alerting and spacing.
Congruency effects (differences in RTs between incongruent and congruent trials) are shown as a function of alerting and spacing in Fig. 2. Congruency effects were larger on alert trials (65 ms) than on no-alert trials (56 ms), yielding a significant interaction between congruency and alerting. Congruency effects decreased from 79 ms to 41 ms as stimulus spacing increased from narrow to wide, yielding a significant interaction between congruency and spacing. Note that the diminished congruency effect with wide spacing was still significant, t(110) = 19.98, p < .001, d = 1.90.

Congruency effects on response times as a function of alerting and spacing in Experiment 1. Error bars represent standard errors of the means
Congruency effects were also influenced by the combination of alerting and spacing, yielding a significant three-way interaction (see Table 2 and Fig. 2). I conducted Bayes-factor analyses using scaled JZS Bayes factors with r = 1 (Rouder, Speckman, Sun, Morey, & Iverson, 2009) to dissect this interaction. For narrowly spaced stimuli, the congruency effect was 16 ms larger on alert than on no-alert trials, and a difference in congruency effects was 2,682.6 times more likely than was no difference. In contrast, for widely spaced stimuli, the congruency effect was only 1 ms larger on alert than on no-alert trials, and no difference in congruency effects was 12.2 times more likely than was a difference. An analysis of the three-way interaction—assessing whether modulation of the congruency effect by alerting varied with spacing—indicated that a difference was 40.3 times more likely than was no difference.
Discussion
The results of Experiment 1 have implications for the diffuse attention hypothesis. Replicating previous research involving variants of the ANT (e.g., Fan et al., 2002; MacLeod et al., 2010; Schneider, 2018), alert trials yielded shorter overall RTs but larger congruency effects than did no-alert trials. Consistent with previous research involving the flanker task (e.g., B. A. Eriksen & Eriksen, 1974; Miller, 1991), congruency effects decreased with wider spacing of stimuli. Critically, there was a significant three-way interaction between congruency, alerting, and spacing in RTs, but the form of the interaction differed from that predicted on the basis of the diffuse attention hypothesis. Instead of the difference in congruency effects between alert and no-alert trials being larger for wide than for narrow spacing, the difference was robust for narrow spacing but eliminated for wide spacing (see Fig. 2).
The obtained three-way interaction is difficult to reconcile with the diffuse attention hypothesis. It is unclear why a wider attentional spotlight triggered by alerting cues would benefit overall RTs when stimuli are widely spaced but would not increase distractor influence. One might argue that the most distant distractors were too far away to fall within even a widened spotlight, but the original prediction still holds if attention were distributed across the same number of distractors on alert and no-alert trials, and the attentional capacity allocated to distractors varied with spotlight size (Heitz & Engle, 2007; White et al., 2011).
The data from Experiment 1 are also challenging for the alternative hypotheses outlined earlier for the interaction between congruency and alerting. The hypothesis that increased alertness inhibits cognitive control (e.g., Callejas et al., 2004) cannot explain why inhibition would occur with narrow spacing but not with wide spacing, despite evidence of increased alertness (alerting effect) and impaired cognitive control (congruency effect) with wide spacing. The hypothesis that alerting cues modulate response selection by activating established stimulus–response associations (e.g., Fischer et al., 2012) can explain the data in a post hoc manner only if one makes the questionable assumption that narrowly spaced stimuli directly activated associated responses but widely spaced stimuli did not. The early onset hypothesis (Nieuwenhuis & de Kleijn, 2013) involves the assumption that attention is more diffuse when response selection starts on alert than on no-alert trials, but the data from Experiment 1 do not support that assumption.
Regarding the global processing hypothesis, the original idea (Weinbach & Henik, 2011) was that alerting cues produce a bias toward global processing of stimuli, improving stimulus detection but impairing target–distractor discrimination. Weinbach and Henik (2012) elaborated this idea by suggesting that alerting cues increase the accessibility of spatial information by expanding the attentional spotlight (i.e., the diffuse attention hypothesis). The data from Experiment 1 are inconsistent with this suggestion, as are the data from Seibold (2018), but it might be possible to retain the global processing hypothesis in modified form if alertness affects how stimuli are processed within the spotlight rather than the size of the spotlight.
The results of Experiment 1 led me to consider a variant of the global processing hypothesis—which I call the perceptual grouping hypothesis—whereby alerting cues elicit a global processing bias by increasing the probability that stimuli are grouped into the same perceptual object (e.g., a row of five arrows) rather than represented as multiple distinct objects (e.g., five separate arrows). On the one hand, perceptually grouping the stimuli could allow attention to be directed quickly to a single object, which might be an adaptive aspect of increased alertness that yields the alerting effect on overall RTs. On the other hand, perceptually grouping the stimuli could make it harder to distinguish between target and distractor features (Logan, 1996), which would facilitate and impair response selection for congruent and incongruent stimuli, respectively, yielding a larger congruency effect on alert than on no-alert trials.
The perceptual grouping hypothesis can explain the data in Experiment 1 if stimulus spacing affects the baseline probability of grouping the stimuli. Drawing on the Gestalt law of perceptual organization by proximity (for overviews, see Hartmann, 1935; Wagemans et al., 2012), perceptual grouping might be more likely with narrower spacing of stimuli. If it is harder to distinguish between target and distractor features when stimuli are grouped, then one would expect larger congruency effects with narrower spacing, as found in Experiment 1. If increased alertness promotes perceptual grouping, then its effect on performance might be modulated by the baseline probability of grouping, as determined by stimulus proximity. For narrowly spaced stimuli that are likely to be grouped even on no-alert trials, increased alertness might promote more grouping, resulting in a larger congruency effect on alert than on no-alert trials. For widely spaced stimuli with a low baseline probability of perceptual grouping, increased alertness might have a negligible effect, resulting in little or no difference in congruency effects between alert and no-alert trials.
If perceptual grouping is the key to the interaction between congruency and alerting, then other grouping manipulations would be expected to modulate the interaction in much the same way that stimulus spacing did in Experiment 1. According to the Gestalt law of perceptual organization by similarity (Hartmann, 1935; Wagemans et al., 2012), stimuli with similar physical properties (e.g., same color) tend to be grouped. In the flanker task, congruency effects are larger when targets and distractors are perceptually grouped on the basis of similarity than when they are not (Baylis & Driver, 1992; Harms & Bundesen, 1983; Kramer & Jacobson, 1991; Luo & Proctor, 2016). The main explanation of this finding is that dissimilar stimuli (e.g., targets and distractors in different colors) are easier to visually parse than are similar stimuli, improving selective attention to the target and reducing distractor influence.
In Experiment 2, I manipulated the colors of targets and distractors to test the perceptual grouping hypothesis. The method was similar to that of Experiment 1, except the arrow stimuli were narrowly spaced and varied in color grouping (see Fig. 1). On same-color trials, the target and distractor arrows were displayed in the same color (either light blue or yellow). On different-color trials, the target arrow was displayed in one color (e.g., light blue) and the distractor arrows were displayed in the alternate color (e.g., yellow). Perceptual grouping based on similarity would suggest more grouping of targets and distractors on same-color than on different-color trials.
I expected shorter overall RTs but larger congruency effects on alert than on no-alert trials, as in Experiment 1 and in previous studies (e.g., Fan et al., 2002; MacLeod et al., 2010; Schneider, 2018). I also expected congruency effects to be smaller for different-color trials than for same-color trials (e.g., as in Baylis & Driver, 1992; Kramer & Jacobson, 1991). Critically, the perceptual grouping hypothesis leads to the prediction of a three-way interaction between congruency, alerting, and color grouping, analogous to the three-way interaction involving stimulus spacing that was obtained in Experiment 1. More specifically, the prediction is for a smaller difference in congruency effects between alert and no-alert trials on different-color trials than on same-color trials. The occurrence of this three-way interaction would support the perceptual grouping hypothesis.Footnote 1
Experiment 2
Method
Subjects
A total of 110 students from Purdue University participated for course credit. None of them had participated in Experiment 1. A simulation-based power analysis using the data from Experiment 1 indicated that 110 subjects would provide 85% power to detect a three-way interaction between congruency, alerting, and color grouping that was the same magnitude as the corresponding interaction involving stimulus spacing that was observed in Experiment 1. Data from one additional subject were excluded for a mean RT (averaged over all conditions) more than four standard deviations above the group mean. All subjects in Experiments 2–4 reported having normal or corrected-to-normal vision and normal color vision.
Apparatus, task, and stimuli
These aspects of the experiment were identical to those of Experiment 1, except the arrow stimuli were narrowly spaced and varied in color grouping. The central fixation cross and alerting cues were always displayed in white (the background was black), but the arrows were light blue or yellow. Target and distractor arrows were displayed in the same color or in different colors on each trial (see Fig. 1). Whether the target arrow was light blue or yellow varied randomly across trials.
Procedure
The procedure was identical to that of Experiment 1, except for the following changes. During the instructions, subjects performed eight example no-alert trials (reflecting all combinations of congruency, color grouping, and target direction) with accuracy feedback. Afterward, they performed 13 blocks of 32 trials per block without accuracy feedback. Every possible combination of alerting, congruency, color grouping, target color, and target direction occurred equally often in random order in each block. The only change to the trial procedure was that the randomly selected fixation interval was 1,000; 1,300; 1,600; or 1,900 ms; the intervals occurred equally often in each block.
Results
Data trimming was identical to that of Experiment 1 (1.7% of trials were RT outliers). Mean RTs and error rates are reported as a function of congruency (incongruent or congruent), alerting (no alert or alert), and color grouping (same or different) in Table 3. The results of repeated-measures ANOVAs with those variables as factors are summarized in Table 4. The mean error rate was 2.2%, and there were no indications of speed–accuracy trade-offs; therefore, I focused on the RT data.
Performance was worse for incongruent stimuli (523 ms) than for congruent stimuli (457 ms), yielding a significant main effect of congruency. Subjects responded faster on alert trials (477 ms) than on no-alert trials (503 ms), yielding a significant main effect of alerting. Subjects were also faster on different-color trials (474 ms) than on same-color trials (507 ms), yielding a significant main effect of color grouping.
Congruency effects are shown as a function of alerting and color grouping in Fig. 3. Congruency effects were larger on alert trials (73 ms) than on no-alert trials (59 ms), yielding a significant interaction between congruency and alerting. Congruency effects were smaller for different-color trials (50 ms) than for same-color trials (82 ms), yielding a significant interaction between congruency and color grouping.

Congruency effects on response times as a function of alerting and color grouping in Experiment 2. Error bars represent standard errors of the means
The three-way interaction between congruency, alerting, and color grouping was nonsignificant (see Table 4 and Fig. 3). I conducted Bayes-factor analyses analogous to those done for Experiment 1. For same-color trials, the congruency effect was 15 ms larger on alert than on no-alert trials, and a difference in congruency effects was 38.9 times more likely than was no difference. For different-color trials, the congruency effect was 14 ms larger on alert than on no-alert trials, and a difference in congruency effects was 17,788.1 times more likely than was no difference. An analysis of the three-way interaction—assessing whether modulation of the congruency effect by alerting varied with color grouping—indicated that no difference was 12.7 times more likely than was a difference.Footnote 2
Discussion
The results of Experiment 2 have implications for the perceptual grouping hypothesis. Replicating previous research involving variants of the ANT (e.g., Fan et al., 2002; MacLeod et al., 2010; Schneider, 2018) and Experiment 1, alert trials yielded shorter overall RTs but larger congruency effects than did no-alert trials. Consistent with previous research involving the flanker task (e.g., Baylis & Driver, 1992; Kramer & Jacobson, 1991), congruency effects were smaller for different-color trials than for same-color trials. Critically, the three-way interaction between congruency, alerting, and color grouping was nonsignificant in RTs (supported by a Bayes-factor analysis that provided strong evidence for a null interaction), contrary to the prediction based on the perceptual grouping hypothesis.
The absence of a three-way interaction is difficult to reconcile with the perceptual grouping hypothesis, at least in its general form. If increased alertness promotes perceptual grouping, then manipulations that lower the probability of grouping the stimuli should counter that effect and reduce the difference in congruency effects between alert and no-alert trials. The relevant interaction was obtained when manipulating perceptual grouping by proximity in Experiment 1, but not when manipulating perceptual grouping by similarity (based on color) in Experiment 2 (compare Figs. 2 and 3). Before interpreting these results further, it seemed prudent to try to replicate the major findings.
In Experiment 3, I manipulated the colors of targets and distractors to test the perceptual grouping hypothesis again. The method was nearly identical to that of Experiment 2, except the arrow stimuli were always displayed in black (matching the screen background), and I varied the color grouping of filled rectangles in which the arrows were displayed (see Fig. 1). On same-color trials, the target and distractor arrows were displayed inside rectangles of the same color (either light blue or yellow). On different-color trials, the target arrow was displayed inside a rectangle of one color (e.g., light blue) and the distractor arrows were displayed inside rectangles of the alternate color (e.g., yellow). Perceptual grouping based on similarity would suggest more grouping of targets and distractors on same-color than on different-color trials, leading to the same predicted three-way interaction that was investigated in Experiment 2.
Experiment 3
Method
Subjects
A total of 110 students from Purdue University participated for course credit. None of them had participated in Experiments 1 or 2. No subjects had either excessively long mean RTs or mean error rates greater than 20%.
Apparatus, task, stimuli, and procedure
These aspects of the experiment were identical to those of Experiment 2, except the arrow stimuli were displayed in black and inside of filled rectangles that were light blue or yellow. Target and distractor arrows were displayed inside rectangles of the same color or different colors on each trial (see Fig. 1). Whether the rectangle containing the target arrow was light blue or yellow varied randomly across trials.
Results
Data trimming was identical to that of Experiments 1 and 2 (1.6% of trials were RT outliers). Mean RTs and error rates are reported as a function of congruency (incongruent or congruent), alerting (no alert or alert), and color grouping (same or different) in Table 5. The results of repeated-measures ANOVAs with those variables as factors are summarized in Table 6. The mean error rate was 2.8%, and there were no indications of speed–accuracy trade-offs; therefore, I focused on the RT data.
Performance was worse for incongruent stimuli (517 ms) than for congruent stimuli (458 ms), yielding a significant main effect of congruency. Subjects responded faster on alert trials (475 ms) than on no-alert trials (500 ms), yielding a significant main effect of alerting. Subjects were also faster on different-color trials (476 ms) than on same-color trials (500 ms), yielding a significant main effect of color grouping. Alerting effects were smaller on different-color trials (23 ms) than on same-color trials (27 ms), yielding a significant interaction between alerting and color grouping.
Congruency effects are shown as a function of alerting and color grouping in Fig. 4. Congruency effects were larger on alert trials (62 ms) than on no-alert trials (57 ms), yielding a significant interaction between congruency and alerting. Congruency effects were smaller for different-color trials (47 ms) than for same-color trials (72 ms), yielding a significant interaction between congruency and color grouping.

Congruency effects on response times as a function of alerting and color grouping in Experiment 3. Error bars represent standard errors of the means
The three-way interaction between congruency, alerting, and color grouping was nonsignificant (see Table 6 and Fig. 4). I conducted Bayes-factor analyses identical to those done for Experiment 2. For same-color trials, the congruency effect was 4 ms larger on alert than on no-alert trials, but no difference in congruency effects was 5.9 times more likely than was a difference. For different-color trials, the congruency effect was 6 ms larger on alert than on no-alert trials, but no difference in congruency effects was 1.4 times more likely than was a difference. An analysis of the three-way interaction—assessing whether modulation of the congruency effect by alerting varied with color grouping—indicated that no difference was 12.1 times more likely than was a difference.
Discussion
The results of Experiment 3 have modest implications for the perceptual grouping hypothesis because of the unexpectedly weak interaction between congruency and alerting. Replicating previous research involving variants of the ANT (e.g., Fan et al., 2002; MacLeod et al., 2010; Schneider, 2018) and Experiments 1 and 2, alert trials yielded shorter overall RTs but larger congruency effects than did no-alert trials. However, the difference in congruency effects between alert and no-alert trials was only 5 ms, which was statistically significant in the ANOVA, but not supported by Bayes factors favoring a difference. Consistent with previous research involving the flanker task (e.g., Baylis & Driver, 1992; Kramer & Jacobson, 1991) and Experiment 2, congruency effects were smaller for different-color trials than for same-color trials. Critically, the three-way interaction between congruency, alerting, and color grouping was nonsignificant in RTs (supported by a Bayes-factor analysis that provided strong evidence for a null interaction), consistent with the data from Experiment 2, and contrary to the prediction based on the perceptual grouping hypothesis.
It is unclear why changing the nature of the color-grouping manipulation from that used in Experiment 2 resulted in a weaker interaction between congruency and alerting in Experiment 3. Nevertheless, the overall data patterns were similar in Experiments 2 and 3 (compare Figs. 3 and 4), with replication of the key finding of no three-way interaction between congruency, alerting, and color grouping. Thus, neither data set supports the perceptual grouping hypothesis. Considering that the most compelling evidence against the diffuse attention and perceptual grouping hypotheses is from Experiments 1 and 2, respectively, it seemed worthwhile to investigate whether the two qualitatively distinct data patterns (see Figs. 2 and 3) could be replicated in the same context.
In Experiment 4, I manipulated stimulus spacing and the colors of targets and distractors in a factorial design for a combined test of the diffuse attention and perceptual grouping hypotheses. Stimulus spacing was manipulated in the same way as in Experiment 1: The spacing of adjacent arrows was either narrow or wide. Color grouping was manipulated in the same way as in Experiment 2: The target and distractor arrows were displayed either in the same color or in different colors. The factorial combination of these two manipulations resulted in four basic types of stimulus displays (see Fig. 1). Two three-way interactions were of particular interest. First, if spacing affects performance as it did in Experiment 1, then the difference in congruency effects between alert and no-alert trials should be larger for narrow spacing than for wide spacing, resulting in a three-way interaction between congruency, alerting, and spacing. Second, if color grouping affects performance as it did in Experiment 2, then the difference in congruency effects between alert and no-alert trials should be similar for same-color and different-color trials, resulting in no three-way interaction between congruency, alerting, and color grouping. The occurrence of both data patterns in the same context would bolster the results of Experiments 1 and 2.
Experiment 4
Method
Subjects
A total of 111 students from Purdue University participated for course credit. None of them had participated in Experiments 1, 2, or 3. The targeted sample size of 110 subjects was exceeded because an extra subject was available on the final day of data collection. Data from five additional subjects were excluded: Three of those subjects had mean RTs (averaged over all conditions) more than four standard deviations above the group mean; the other two subjects had mean error rates (averaged over all conditions) greater than 20%.
Apparatus, task, and stimuli
These aspects of the experiment were identical to those of Experiments 1 and 2, except for the factorial combination of stimulus spacing and color grouping. The central fixation cross and alerting cues were always displayed in white (the background was black), but the arrows were light blue or yellow. The spacing of adjacent arrows (edge-to-edge distance) was either narrow (0.11°) or wide (1.26°), as in Experiment 1, and the target and distractor arrows were displayed in the same color or in different colors on each trial, as in Experiment 2 (see Fig. 1).
Procedure
The procedure was identical to that of Experiment 2, except for the following changes. During the instructions, subjects did eight example no-alert trials (reflecting all combinations of congruency, spacing, and color grouping) with accuracy feedback. Afterward, they performed 15 blocks of 64 trials per block without accuracy feedback. Every possible combination of alerting, congruency, spacing, color grouping, target color, and target direction occurred equally often in random order in each block.
Results
Data trimming was identical to that of Experiments 1, 2, and 3 (1.9% of trials were RT outliers). Mean RTs and error rates are reported as a function of congruency (incongruent or congruent), alerting (no alert or alert), spacing (narrow or wide), and color grouping (same or different) in Table 7. The results of repeated-measures ANOVAs with those variables as factors are summarized in Table 8. The mean error rate was 2.9%, and there were no indications of speed–accuracy trade-offs; therefore, I focused on the RT data.
Performance was worse for incongruent stimuli (510 ms) than for congruent stimuli (457 ms), yielding a significant main effect of congruency. Subjects responded faster on alert trials (465 ms) than on no-alert trials (501 ms), yielding a significant main effect of alerting. Subjects were faster when stimulus spacing was wide (468 ms) than when it was narrow (499 ms), yielding a significant main effect of spacing. Subjects were also faster on different-color trials (471 ms) than on same-color trials (496 ms), yielding a significant main effect of color grouping. Alerting effects increased from 32 ms to 40 ms with wider spacing, as in Experiment 1, yielding a significant interaction between alerting and spacing. Alerting effects were smaller on different-color trials (33 ms) than on same-color trials (39 ms), as in Experiment 3, yielding a significant interaction between alerting and color grouping. Color-grouping effects (differences in RTs between same-color and different-color trials) decreased from 36 ms to 16 ms with wider spacing, yielding a significant interaction between spacing and color grouping.
Congruency effects are shown as a function of alerting, spacing, and color grouping in Fig. 5. Congruency effects were larger on alert trials (58 ms) than on no-alert trials (48 ms), yielding a significant interaction between congruency and alerting. Congruency effects decreased from 71 ms to 35 ms as stimulus spacing increased from narrow to wide, yielding a significant interaction between congruency and spacing. Congruency effects were smaller for different-color trials (41 ms) than for same-color trials (65 ms), yielding a significant interaction between congruency and color grouping. Higher order interactions involving congruency were of particular interest, and I conducted accompanying Bayes-factor analyses similar to those done for Experiments 1 and 2.

Congruency effects on response times as a function of alerting, spacing, and color grouping in Experiment 4. Error bars represent standard errors of the means
The three-way interaction between congruency, alerting, and spacing was significant (see Table 8 and Fig. 5), as in Experiment 1. To facilitate comparison with the results of Experiment 1, in which the arrow stimuli were always displayed in the same color, the following numbers in parentheses are from analyses restricted to same-color trials. For narrowly spaced stimuli, the congruency effect was 17 ms (18 ms) larger on alert than on no-alert trials, and a difference in congruency effects was 606.4 times (44.6 times) more likely than was no difference. In contrast, for widely spaced stimuli, the congruency effect was only 3 ms (−1 ms) larger on alert than on no-alert trials, and no difference in congruency effects was 5.8 times (13.0 times) more likely than was a difference. An analysis of the three-way interaction—assessing whether modulation of the congruency effect by alerting varied with spacing—indicated that a difference was 56.1 times (16.6 times) more likely than was no difference.
In contrast, the three-way interaction between congruency, alerting, and color grouping was nonsignificant (see Table 8 and Fig. 5), as in Experiment 2. To facilitate comparison with the results of Experiment 2, in which the arrow stimuli were always narrowly spaced, the following numbers in parentheses are from analyses restricted to narrowly spaced stimuli. For same-color trials, the congruency effect was 9 ms (18 ms) larger on alert than on no-alert trials, and a difference in congruency effects was 3.0 times (44.6 times) more likely than was no difference. For different-color trials, the congruency effect was 11 ms (16 ms) larger on alert than on no-alert trials, and a difference in congruency effects was 12.9 times (23.6 times) more likely than was no difference. An analysis of the three-way interaction—assessing whether modulation of the congruency effect by alerting varied with color grouping—indicated that no difference was 11.1 times (12.2 times) more likely than was a difference.
The only remaining significant effect for RTs was a three-way interaction between congruency, spacing, and color grouping (see Table 8). For same-color trials, the congruency effect was 50 ms larger for narrow than for wide spacing, and a difference in congruency effects was 4.9 × 1019 times more likely than was no difference. For different-color trials, the congruency effect was 22 ms larger for narrow than for wide spacing, and a difference in congruency effects was 1.7 × 109 times more likely than was no difference. An analysis of the three-way interaction—assessing whether modulation of the congruency effect by spacing varied with color grouping—indicated that a difference was 374,322.3 times more likely than was no difference.
Discussion
The results of Experiment 4 have implications for the diffuse attention and perceptual grouping hypotheses. Replicating previous research involving variants of the ANT (e.g., Fan et al., 2002; MacLeod et al., 2010; Schneider, 2018) and Experiments 1–3, alert trials yielded shorter overall RTs but larger congruency effects than did no-alert trials. Consistent with previous research involving the flanker task (e.g., B. A. Eriksen & Eriksen, 1974; Miller, 1991) and Experiment 1, congruency effects decreased with wider spacing of stimuli. Also consistent with previous research involving the flanker task (e.g., Baylis & Driver, 1992; Kramer & Jacobson, 1991), as well as Experiments 2 and 3, congruency effects were smaller for different-color trials than for same-color trials.
There were two critical findings involving three-way interactions. First, the three-way interaction between congruency, alerting, and spacing was significant, reflecting a reliable difference in congruency effects between alert and no-alert trials for narrow spacing, but no difference for wide spacing. This data pattern replicates Experiment 1 (compare Fig. 2 with the same-color data in Fig. 5) and is inconsistent with the diffuse attention hypothesis. Second, the three-way interaction between congruency, alerting, and color grouping was nonsignificant, reflecting a difference in congruency effects between alert and no-alert trials that was similar for same-color and different-color trials. This data pattern replicates Experiment 2 (compare Fig. 3 with the narrow-spacing data in Fig. 5) and is inconsistent with the perceptual grouping hypothesis. Thus, the qualitatively distinct data patterns obtained in Experiments 1 and 2 were replicated in the same context in Experiment 4.
General discussion
A puzzling interaction involving alertness and cognitive control is indicated by the finding of faster performance but larger congruency effects on alert trials than on no-alert trials in selective attention tasks (e.g., MacLeod et al., 2010; Nieuwenhuis & de Kleijn, 2013; Schneider, 2018). One explanation for this finding is the diffuse attention hypothesis (McConnell & Shore, 2011; Weinbach & Henik, 2012), which is the idea that alerting cues trigger a widened attentional spotlight that improves stimulus detection but increases distractor processing. In Experiment 1 (and again in Experiment 4), I tested the diffuse attention hypothesis by manipulating stimulus spacing in a variant of the ANT (Fan et al., 2002).
Replicating previous research, alert trials yielded shorter overall RTs but larger congruency effects than did no-alert trials (Fan et al., 2002; MacLeod et al., 2010; Schneider, 2018), and congruency effects decreased with wider spacing of stimuli (B. A. Eriksen & Eriksen, 1974; Miller, 1991). Critically, there was a significant three-way interaction between congruency, alerting, and spacing in RTs, but the form of the interaction differed from that predicted on the basis of the diffuse attention hypothesis. Instead of the difference in congruency effects between alert and no-alert trials being larger for wide than for narrow spacing, the difference was robust for narrow spacing but eliminated for wide spacing (see Figs. 2 and 5). These data suggest that the diffuse attention hypothesis is not a viable explanation of the interaction between congruency and alerting, which is the same conclusion reached by Seibold (2018) based on probe-RT data. Thus, there is now converging evidence that increased alertness does not seem to be associated with a widened attentional spotlight.
The results of Experiment 1 led me to propose a variant of the global processing hypothesis (Weinbach & Henik, 2011, 2012) called the perceptual grouping hypothesis, which is the idea that alerting cues elicit a global processing bias by increasing the probability that stimuli are grouped into the same perceptual object rather than represented as multiple distinct objects. Drawing on Gestalt laws of perceptual organization, I reasoned that a color-based manipulation of grouping by similarity might yield results analogous to those obtained with the spacing manipulation of grouping by proximity. In Experiments 2–4, I tested the perceptual grouping hypothesis by manipulating the color grouping of targets and distractors in a variant of the ANT.
Replicating previous research, alert trials yielded shorter overall RTs but larger congruency effects than did no-alert trials, and congruency effects were smaller for different-color trials than for same-color trials (Baylis & Driver, 1992; Kramer & Jacobson, 1991). Critically, the three-way interaction between congruency, alerting, and color grouping was nonsignificant in RTs (supported by Bayes-factor analyses that provided strong evidence for null interactions), contrary to the prediction based on the perceptual grouping hypothesis. That is, despite the overall reduction in congruency effects from same-color to different-color trials, the difference in congruency effects between alert and no-alert trials was unaffected (see Figs. 3–5). In Experiment 4, the null interaction between congruency, alerting, and color grouping was obtained in the same context as a significant interaction between congruency, alerting, and spacing. These data suggest that the perceptual grouping hypothesis, at least in its general form, is not a viable explanation of the interaction between congruency and alerting.
Toward a spatial grouping hypothesis
The present data are difficult to reconcile with any of the existing hypotheses in the literature, motivating consideration of alternative ideas about how alertness is related to cognitive control. There are at least three lines of evidence that point to the importance of spatial attention. First, the present study showed that the interaction between congruency and alerting was modulated by a spatial variable (stimulus spacing) and not by a nonspatial variable (color grouping), even though both variables affected overall congruency effects. Second, the interaction between congruency and alerting has been obtained in tasks with a spatial attention component, such as the flanker task (or ANT; e.g., Fan et al., 2002; MacLeod et al., 2010; Redick & Engle, 2006; Schneider, 2018) and the Simon task (e.g., Böckler et al., 2011; Fischer et al., 2010; Klein & Ivanoff, 2011; Soutschek, Müller, & Schubert, 2013, Experiment 1), but not in tasks without a spatial component, such as the Stroop task (Weinbach & Henik, 2012, Experiment 2; Soutschek et al., 2013, Experiment 2). Third, task-switching studies have revealed that switch costs (worse performance for task switches than for task repetitions), which are routinely interpreted as indices of cognitive control, are modulated by alertness when switching between spatial tasks (Meiran & Chorev, 2005), but not when switching between nonspatial tasks (Schneider, 2017).
These findings suggest that spatial characteristics of attention underlie the interaction between congruency and alerting (Weinbach & Henik, 2012). For this reason, I propose a refinement of the perceptual grouping hypothesis—which I call the spatial grouping hypothesis—that emphasizes spatial attention. The spatial grouping hypothesis is based on the idea that spatial attention mechanisms determine how displays of multiple stimuli arrayed in space are parsed into distinct object representations used in subsequent processing associated with stimulus categorization and response selection. The main assumption is that increased alertness leads to more spatial grouping of stimuli, which is more specific than perceptual grouping because the latter can involve nonspatial stimulus dimensions. In selective attention tasks, spatially grouping the stimuli could make it harder to distinguish between targets and distractors, facilitating and impairing response selection for congruent and incongruent stimuli, respectively. The result would be a larger congruency effect on alert than on no-alert trials.
One way of linking alertness to spatial grouping would be to modulate the grouping threshold that controls how stimulus displays are parsed into distinct objects. This idea can be expressed in the context of Logan’s (1996) CODE theory of visual attention (CTVA). According to CTVA, a visual stimulus is represented as a distribution of features in space that is centered on the stimulus location, as illustrated in Fig. 6 for an array of arrow stimuli. Summation of feature distributions across stimuli produces a combined distribution called a CODE surface (solid black line in Fig. 6) to which a grouping threshold (gray line) is applied.Footnote 3 Above-threshold regions of the CODE surface are treated as distinct objects, but they can include features from multiple stimuli (e.g., targets and distractors) due to feature distribution overlap. Attention works by sampling features within a chosen above-threshold region, and the sample of features is used as input to stimulus categorization and response selection processes.

Illustration of feature distributions (dashed black lines) and CODE surface (solid black line) for arrow stimuli as a function of spacing in the horizontal dimension. High and low thresholds (thick and thin gray lines, respectively) applied to the CODE surface parse the stimulus display into spatial groups
The grouping threshold plays an important role in CTVA. A high threshold applied to the CODE surface reduces the features available to be sampled because above-threshold regions are smaller, but it improves stimulus discriminability (see Fig. 5 in Logan, 1996) because sampled features are more likely to belong to one particular stimulus. Moreover, a high threshold makes it less likely that stimuli will be grouped in the same region (i.e., as part of the same object). In contrast, a low threshold increases grouping of stimuli but allows for sampling of more features, which could be interpreted as a type of global processing bias (cf. Weinbach & Henik, 2011).
Logan (1996) applied CTVA to an experiment by B. A. Eriksen and Eriksen (1974) involving a flanker task with manipulation of target–distractor spacing. CTVA produced congruency effects that decreased with wider spacing, as well as shorter overall RTs with wider spacing. It produced congruency effects because distractor features were sometimes sampled with target features from above-threshold regions, impairing stimulus categorization when distractors were incongruent. Spacing effects arose from differences in feature distribution overlap and changes in threshold. For narrow spacing, there was considerable overlap among target and distractor feature distributions, necessitating a high threshold to reduce grouping and to increase sampling of target features relative to distractor features (see Fig. 15 in Logan, 1996). For wide spacing, there was less overlap among feature distributions, allowing the threshold to be lowered (without increasing grouping) and improving stimulus discriminability because fewer distractor features were sampled from the above-threshold region centered on the target.
Applying CTVA to the present study, I propose that increased alertness is associated with a lowered threshold for spatial grouping of stimuli. Figure 6 illustrates how linking alertness to the threshold can affect spatial grouping.Footnote 4 The threshold (gray line) is assumed to vary as an inverse function of alertness: as alertness increases, the threshold decreases. Each panel of Fig. 6 shows the same high and low thresholds (thick and thin gray lines, respectively), which might reflect small differences in alertness on alert and no-alert trials, respectively. What differs between panels is stimulus spacing (narrow or wide), which affects the resulting CODE surface and how the threshold parses it into objects.
For narrowly spaced stimuli (top panel of Fig. 6), the high threshold (thick gray line) parses the CODE surface into three above-threshold regions: two small regions for the outer distractors and a large region that includes the target and inner distractors. When the large region is attended, features sampled from it can come from the target and inner distractors, impairing and facilitating subsequent processing on incongruent and congruent trials, respectively, resulting in a congruency effect. If increased alertness is associated with a slightly lower threshold (thin gray line), then more spatial grouping is possible; in this case, there is now a single above-threshold region that includes the target and all the distractors. There is a higher probability of sampling distractor features from that region, resulting in a larger congruency effect than that associated with the higher threshold and lower alertness.
For widely spaced stimuli (bottom panel of Fig. 6), lowering the threshold by the same amount has no effect on the spatial grouping of stimuli. The high threshold (thick gray line) parses the CODE surface into three above-threshold regions: one for each inner distractor and one for the target. When the target region is attended, features sampled from it are more likely to come from the target than from the distractors because of the limited overlap between target and distractor feature distributions, resulting in a smaller congruency effect than that obtained with the same high threshold under narrow spacing. If increased alertness is associated with a slightly lower threshold (thin gray line), spatial grouping does not change: the same three qualitatively distinct above-threshold regions exist. Feature sampling would be relatively unaffected (i.e., most of the features sampled from the target region would still come from the target), resulting in a small congruency effect of similar magnitude to that obtained with the high threshold.
The net result of lowering the threshold when alertness is increased would be a larger congruency effect on alert than on no-alert trials, primarily when stimuli are narrowly spaced and already likely to be spatially grouped. As stimulus spacing increases, the congruency effect would decrease because of less overlap between target and distractor feature distributions, as well as less grouping of target and distractors in the same above-threshold region. Lowering the threshold under wide spacing might lead to little or no change in spatial grouping and, consequently, little or no difference in congruency effects between alert and no-alert trials. This interpretation of the spatial grouping hypothesis in the context of CTVA seems consistent with the spacing data in Experiments 1 and 4, and with the color-grouping data in Experiments 2–4. Color grouping is not a spatial variable, so it would affect neither the spatial distributions of features nor the grouping threshold that parses the CODE surface into spatially defined objects. However, CTVA defines objects based only on their locations, which is a limitation of the original theory. Objects can be defined along other dimensions that affect performance in selective attention tasks, as demonstrated by the color-grouping effects in the present study. Logan (1996) discussed how other aspects of the theory, such as its similarity parameters (see Bundesen, 1990), might allow it to address grouping by similarity based on shape or other dimensions.
I want to emphasize that changing the grouping threshold in CTVA need not be the only way in which alertness might affect stimulus processing. Modeling work in other contexts suggests that increased alertness is associated with faster visual processing speed or a lower perceptual threshold for encoding stimuli into visual short-term memory (Matthias et al., 2010; Petersen et al., 2017). Bundesen, Vangkilde, and Habekost (2015) proposed that modeling of visual bias might benefit from including a parameter representing the general level of alertness. As the value of the alertness parameter increases, the speed with which a stimulus is categorized (i.e., encoded into visual short-term memory) also increases. The original formulation of CTVA lacks explicit parameters or mechanisms associated with alertness, so the proposal of Bundesen and colleagues might be a way of extending the theory. However, modeling alertness and its effects on processing in selective attention tasks is beyond the scope of the present study.
Conclusion
Linking alertness to the grouping threshold in CTVA is one approach to accounting for the present data. It provides a mechanistic basis for the spatial grouping hypothesis and for producing something akin to a global processing bias (Weinbach & Henik, 2011, 2012). However, instead of alertness affecting the size of the attentional spotlight, it might modulate the spatial grouping threshold that influences what is sampled by attention. An objective for future research would be to test whether CTVA or other models of selective attention in the flanker task (e.g., Hübner et al., 2010; White et al., 2011) can be adapted to variants of the ANT to better explain how alertness is related to cognitive control in the human attention system.
Author note
I thank Timothy Durbin, Sammi Franklin, Carli Hoover, Shreya Kanchan, Michael Kanczuzewski, Kelsey Lynch, Yijie Peng, Tessa Stephens, Sami Wagner, and Ashley Young for assistance with data collection. I also thank Gordon Logan, Colin MacLeod, Noam Weinbach, and an anonymous reviewer for comments on previous drafts of the manuscript.
Notes
Experiments 2 and 3 were preregistered tests of the perceptual grouping hypothesis. The protocol is publicly available at https://aspredicted.org/xj9tx.pdf (Note that Experiments 2 and 3 are referred to as Experiments 1 and 2, respectively, in the protocol; the present Experiment 1 is referenced as an unpublished experiment.) Although Experiments 1 and 4 were not preregistered, they conformed to many of the details in the protocol, including subject/trial exclusion criteria and analytical methods.
There was a notable discrepancy between the RT and error data in Experiment 2: There was a significant three-way interaction between congruency, alerting, and color grouping for error rates (see Table 4), reflecting a larger difference in congruency effects between alert and no-alert trials for different-color trials (1.7%) than for same-color trials (0.4%)—a pattern that is opposite the prediction based on the perceptual grouping hypothesis. However, a Bayes-factor analysis for the three-way interaction indicated that a difference was only 1.1 times more likely than was no difference, and the corresponding interactions were nonsignificant in Experiments 3 and 4; therefore, the effect might be spurious.
The threshold for spatial grouping of stimuli in CTVA is qualitatively distinct from “thresholds” in other contexts. For example, Petersen, Petersen, Bundesen, Vangkilde, and Habekost (2017) found that increased alertness was associated with a lower perceptual threshold (a temporal parameter) for encoding stimuli into visual short-term memory during a short-exposure letter-recognition task.
Figure 6 illustrates how different thresholds can result in different spatial groupings of stimuli under narrow and wide spacing conditions in CTVA at a conceptual level. It does not reflect any model-based estimation of feature distributions or thresholds. In addition, the “high” and “low” threshold labels used in the text designate the levels of the thresholds relative to each other, not in absolute terms.
References
Baylis, G. C., & Driver, J. (1992). Visual parsing and response competition: The effect of grouping factors. Perception & Psychophysics, 51, 145–162.
Böckler, A., Alpay, G., & Stürmer, B. (2011). Accessory stimuli affect the emergence of conflict, not conflict control: A Simon-task ERP study. Experimental Psychology, 58, 102–109.
Bundesen, C. (1990). A theory of visual attention. Psychological Review, 97, 523–547.
Bundesen, C., Vangkilde, S., & Habekost, T. (2015). Components of visual bias: A multiplicative hypothesis. Annals of the New York Academy of Sciences, 1339, 116–124.
Callejas, A., Lupiáñez, J., Funes, M. J., & Tudela, P. (2005). Modulations among the alerting, orienting, and executive control networks. Experimental Brain Research, 167, 27–37.
Callejas, A., Lupiáñez, J., & Tudela, P. (2004). The three attentional networks: On their independence and interactions. Brain and Cognition, 54, 225–227.
Cave, K. R., & Bichot, N. P. (1999). Visuospatial attention: Beyond a spotlight model. Psychonomic Bulletin & Review, 6, 204–223.
Cohen, J. D., Servan-Schreiber, D., & McClelland, J. L. (1992). A parallel distributed processing approach to automaticity. American Journal of Psychology, 105, 239–269.
Eriksen, B. A., & Eriksen, C. W. (1974). Effects of noise letters upon the identification of a target letter in a nonsearch task. Perception & Psychophysics, 16, 143–149.
Eriksen, C. W., & St. James, J. D. (1986). Visual attention within and around the field of focal attention: A zoom lens model. Perception & Psychophysics, 40, 225–240.
Fan, J., McCandliss, B. D., Sommer, T., Raz, A., & Posner, M. I. (2002). Testing the efficiency and independence of attentional networks. Journal of Cognitive Neuroscience, 14, 340–347.
Fischer, R., Plessow, F., & Kiesel, A. (2010). Auditory warning signals affect mechanisms of response selection: Evidence from a Simon task. Experimental Psychology, 57, 89–97.
Fischer, R., Plessow, F., & Kiesel, A. (2012). The effects of alerting signals in action control: Activation of S–R associations or inhibition of executive control processes? Psychological Research, 76, 317–328.
Harms, L., & Bundesen, C. (1983). Color segregation and selective attention in a nonsearch task. Perception & Psychophysics, 33, 11–19.
Hartmann, G. W. (1935). Gestalt psychology: A survey of facts and principles. New York: Ronald Press.
Heitz, R. P., & Engle, R. W. (2007). Focusing the spotlight: Individual differences in visual attention control. Journal of Experimental Psychology: General, 136, 217–240.
Hübner, R., Steinhauser, M., & Lehle, C. (2010). A dual-stage two-phase model of selective attention. Psychological Review, 117, 759–784.
Klein, R. M., & Ivanoff, J. (2011). The components of visual attention and the ubiquitous Simon effect. Acta Psychologica, 136, 225–234.
Kramer, A. F., & Jacobson, A. (1991). Perceptual organization and focused attention: The role of objects and proximity in visual processing. Perception & Psychophysics, 50, 267–284.
LaBerge, D. (1983). Spatial extent of attention to letters and words. Journal of Experimental Psychology: Human Perception and Performance, 9, 371–379.
LaBerge, D., & Brown, V. (1989). Theory of attentional operations in shape identification. Psychological Review, 96, 101–124.
Logan, G. D. (1996). The CODE theory of visual attention: An integration of space-based and object-based attention. Psychological Review, 103, 603–649.
Luo, C., & Proctor, R. W. (2016). Perceptual grouping of objects occupied by target and flankers affects target-flanker interference. Attention, Perception, & Psychophysics, 78, 251–263.
MacLeod, J. W., Lawrence, M. A., McConnell, M. M., Eskes, G. A., Klein, R. M., & Shore, D. I. (2010). Appraising the ANT: Psychometric and theoretical considerations of the Attention Network Test. Neuropsychology, 24, 637–651.
Matthias, E., Bublak, P., Müller, H. J., Schneider, W. X., Krummenacher, J., & Finke, K. (2010). The influence of alertness on spatial and nonspatial components of visual attention. Journal of Experimental Psychology: Human Perception and Performance, 36, 38–56.
Mattler, U. (2006). Distance and ratio effects in the flanker task are due to different mechanisms. Quarterly Journal of Experimental Psychology, 59, 1745–1763.
McConnell, M. M., & Shore, D. I. (2011). Mixing measures: Testing an assumption of the attention network test. Attention, Perception, & Psychophysics, 73, 1096–1107.
Meiran, N., & Chorev, Z. (2005). Phasic alertness and the residual task-switching cost. Experimental Psychology, 52, 109–124.
Miller, J. (1991). The flanker compatibility effect as a function of visual angle, attentional focus, visual transients, and perceptual load: A search for boundary conditions. Perception & Psychophysics, 49, 270–288.
Navon, D. (1977). Forest before trees: The precedence of global features in visual perception. Cognitive Psychology, 9, 353–383.
Nieuwenhuis, S., & de Kleijn, R. (2013). The impact of alertness on cognitive control. Journal of Experimental Psychology: Human Perception and Performance, 39, 1797–1801.
Petersen, A., Petersen, A. H., Bundesen, C., Vangkilde, S., & Habekost, T. (2017). The effect of phasic auditory alerting on visual perception. Cognition, 165, 73–81.
Posner, M. I. (1980). Orienting of attention. Quarterly Journal of Experimental Psychology, 32, 3–25.
Posner, M. I., & Boies, S. J. (1971). Components of attention. Psychological Review, 78, 391–408.
Posner, M. I., & Petersen, S. E. (1990). The attention system of the human brain. Annual Review of Neuroscience, 13, 25–42.
Redick, T. S., & Engle, R. W. (2006). Working memory capacity and attention network test performance. Applied Cognitive Psychology, 20, 713–721.
Rouder, J. N., Speckman, P. L., Sun, D., Morey, R. D., & Iverson, G. (2009). Bayesian t tests for accepting and rejecting the null hypothesis. Psychonomic Bulletin & Review, 16, 225–237.
Schneider, D. W. (2017). Phasic alertness and residual switch costs in task switching. Journal of Experimental Psychology: Human Perception and Performance, 43, 317–327.
Schneider, D. W. (2018). Alertness and cognitive control: Testing the early onset hypothesis. Journal of Experimental Psychology: Human Perception and Performance. https://doi.org/10.1037/xhp0000497
Seibold, V. C. (2018). Do alerting signals increase the size of the attentional focus? Attention, Perception, & Psychophysics, 80(2), 402–425. https://doi.org/10.3758/s13414-017-1451-1
Soutschek, A., Müller, H. J., & Schubert, T. (2013). Conflict-specific effects of accessory stimuli on cognitive control in the Stroop task and the Simon task. Experimental Psychology, 60, 140–147.
Wagemans, J., Elder, J. H., Kubovy, M., Palmer, S. E., Peterson, M. A., Singh, M., & von der Heydt, R. (2012). A century of Gestalt psychology in visual perception: I. Perceptual grouping and figure–ground organization. Psychological Bulletin, 138, 1172–1217.
Weinbach, N., & Henik, A. (2011). Phasic alertness can modulate executive control by enhancing global processing of visual stimuli. Cognition, 121, 454–458.
Weinbach, N., & Henik, A. (2012). The relationship between alertness and executive control. Journal of Experimental Psychology: Human Perception and Performance, 38, 1530–1540.
Weinbach, N., & Henik, A. (2014). Alerting enhances attentional bias for salient stimuli: Evidence from a global/local processing task. Cognition, 133, 414–419.
White, C. N., Ratcliff, R., & Starns, J. J. (2011). Diffusion models of the flanker task: Discrete versus gradual attentional selection. Cognitive Psychology, 63, 210–238.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Schneider, D.W. Alertness and cognitive control: Toward a spatial grouping hypothesis. Atten Percept Psychophys 80, 913–928 (2018). https://doi.org/10.3758/s13414-018-1491-1
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13414-018-1491-1