
Abstract
In two experiments, we tested whether subliminal abrupt onset cues capture attention in a stimulus-driven way. An onset cue was presented 16 ms prior to the stimulus display that consisted of clearly visible color targets. The onset cue was presented either at the same side as the target (the valid cue condition) or on the opposite side of the target (the invalid cue condition). Because the onset cue was presented 16 ms before other placeholders were presented, the cue was subliminal to the participant. To ensure that this subliminal cue captured attention in a stimulus-driven way, the cue’s features did not match the top-down attentional control settings of the participants: (1) The color of the cue was always different than the color of the non-singleton targets ensuring that a top-down set for a specific color or for a singleton would not match the cue, and (2) colored targets and distractors had the same objective luminance (measured by the colorimeter) and subjective lightness (measured by flicker photometry), preventing a match between the top-down set for target and cue contrast. Even though a match between the cues and top-down settings was prevented, in both experiments, the cues captured attention, with faster response times in valid than invalid cue conditions (Experiments 1 and 2) and faster response times in valid than the neutral conditions (Experiment 2). The results support the conclusion that subliminal cues capture attention in a stimulus-driven way.
Similar content being viewed by others

Evolution has equipped the human visual system with functional mechanisms of attentional guidance to cope with threats imposed on the organism in the course of its phylogeny. Selective attention is the gateway to cognition, allowing us to allocate mental resources to visual input relevant for the task at hand. One very functional attentional guidance mechanism could ensure the failsafe selection of abrupt onsets in the visual periphery. As abrupt onsets in the periphery may contain novel information and possibly a potential threat, shifting attention to the location of an abrupt onset may provide an important survival mechanism (Breitmeyer & Ganz, 1976; Yantis & Jonides, 1984). Because from an ecological viewpoint such a mechanism should operate regardless of the current attentional set, it may not be surprising that abrupt onsets capture attention in an exogenous or stimulus-driven way. The hypothesis that abrupt onsets capture attention in a stimulus-driven way, regardless of top-down control, was initially supported (Jonides, 1981, Posner & Cohen, 1984) where participants searched for a target presented at one of several peripheral locations. If the target happened to be preceded by an abrupt onset cue (the valid cue condition), participants were relatively fast to respond to the target. However, in the invalid cue condition, when the cue was presented at a location other than the target participants were relatively slow (Jonides, 1981, Posner & Cohen, 1984). This validity or cueing effect was assumed to reflect the stimulus-driven capture of attention by the onset cue, which in turn facilitated responding to a target presented at that location relative to a condition in which the target appeared at an uncued location (Posner, 1980). These authors concluded that the validity effect reflected stimulus-driven attentional capture, because the peripheral cues were uninformative about the likely position of the target. Moreover, in Jonides (1981) a validity effect was even found if the participants were asked to actively ignore the cues. On the basis of these latter findings, it was concluded that the cueing effect must be stimulus-driven as participants were not even able to overcome attentional capture when they actively tried.
However, following these initial studies a whole host of new findings questioned the generalizability of the stimulus-driven nature of attentional capture by onsets. Indeed, Folk, Remington, and Johnston (1992) demonstrated that an abrupt onset cue only captured attention if participants searched for an abrupt onset target. If participants searched for a particular color defined target, there was no cueing effect of the abrupt onset cue. This finding led Folk et al. to their top-down contingent-capture theory according to which attentional capture is always contingent on a match between the top-down attentional control settings and the features of the impinging stimuli.
Even though the debate of whether clearly visible (i.e., supraliminal) onset cues capture attention in a stimulus-driven way is still unresolved (see Theeuwes, 2010), more recent work has made the claim that onset cues that are presented subliminally should at least be considered to be stimulus-driven as the observer is not aware that such a cue was presented in the first place (McCormick, 1997; Mulckhuyse, Talsma, & Theeuwes, 2007). A stimulus is considered to be subliminal when the observer is not aware of this stimulus. For example, in Mulckhuyse et al. (2007) one black disk was presented as a single abrupt onset cue in the periphery, either to the right or to the left of the center of a computer screen. Immediately after this cue [with a stimulus onset asynchrony (SOA) of 16 ms], two other black disks were shown as placeholders: one in the center of the screen and one on the opposite side of the cue. In this situation, participants remained unaware that one of the onset cues was presented earlier in time than the other two placeholders as the temporal interval between cue and placeholders was too small to notice this. Even though participants were unaware that the cue had a lead time, it nevertheless captured attention, because participants were faster when a subsequent target was presented at the location of the cue (i.e., valid cue condition) than when it was presented at the location away from the cue (i.e., invalid cue condition). Although it is difficult to ensure absolute subliminality in this kind of experimental protocol (Fuchs & Ansorge, 2012a, 2012b; Fuchs, Theeuwes, & Ansorge, 2013), several findings suggested that the cues were indeed not consciously noticed by the participants, as long as the participants were not directly informed about the presence of the cues. For example, participants were not able to strategically use the subliminal cues while they were able to do so when the cues were presented supraliminally. Also, participants were not aware of the subliminal cues when asked about it.
However, regardless of whether true subliminality can be adequately assessed, for the present discussion there are other reasons to question whether these previous studies indeed measured true stimulus-driven capture. First, in the study by Mulckhuyse et al. (2007), as well as in subsequent research (e.g., Fuchs et al., 2013), the target was always a singleton, implying that it was a unique element among the background of placeholders. For example, the target in Mulckhuyse et al. (2007) was a single small disk. In this situation, in order to find the target, participants may have searched actively for any singleton, implying that the subliminal singleton onset cue could have captured attention in a top-down contingent rather than a stimulus-driven fashion (Bacon & Egeth, 1994; Held, Ansorge, & Müller, 2010; Leber & Egeth, 2006).
So far, only a few experiments tried to rule out this alternative explanation of the subliminal cueing effect. These studies seem to suggest capture that is contingent on a top-down set and argue against true stimulus-driven capture (Fuchs & Ansorge, 2012b). However, even these experiments are not entirely conclusive. Fuchs and Ansorge (2012b), for instance, adapted the subliminal cueing paradigm of Mulckhuyse et al. (2007) and instructed the participants in their Experiment 4 to search for a color target (e.g., blue target) that was presented along with two differently colored distractors (e.g., green and red distractor if the target was blue). The target appeared either on the left or right side while the two other colored distractors appeared at fixation and at the location opposite of that of the target. Under these conditions, participants could not search for a color singleton to locate the target, and in line with a top-down contingent capture explanation a validity effect of the subliminal onset cues was not found. However, this study used a target detection task—that is, participants had to decide whether the target was present or not. Crucially, in the target-absent trials, not only the target was left out but also the distractors. In this situation, the present-absent decision did not require locating the target at all, and the decision could be made on the basis of the presence of both target and distractors. Thus, it is unclear whether the participants had to search for a particular color target or whether they responded to the presence of the whole display of target and distractors. So, for this reason alone the study of Fuchs and Ansorge is inconclusive.
In addition, there is another reason to question whether capture was truly stimulus-driven in these previous studies. In some studies there was a clear target-background luminance contrast (e.g., Fuchs et al., 2013; Mulckhuyse et al, 2007), whereas in other studies the color target was only objectively equated to the luminance of the background (as in Fuchs & Ansorge, 2012b). Because it is well-known that there are individual differences in luminance sensitivity for colors (Gunther & Dobkins, 2002), for some participants the target may have stood out by a stronger contrast than the distractors, even if ever so slightly, which then would allow participants to search for contrasts in a top-down way to find the targets. If this happens, subliminal capture by a cue with a foreground-background contrast could again be the result of top-down contingent capture rather than of stimulus-driven capture.
Experiment 1
Experiment 1 addressed these concerns. First, to rule out top-down singleton search, participants had to search for a color-defined target among two differently colored distractors. For example, the target was blue and the distractors were green and red. In this situation, it is impossible to locate the target as a singleton. In addition, in order to be able to respond to the target, the target needed to be localized. Indeed, participants had to report the orientation of the gap of the Landolt C target which was presented among other Landolt C distractor elements (Fig. 1). Second, to rule out that participants could have used background contrast to find the color target, the subjectively perceived lightness of the target color was carefully equated to that of the distractor colors by individual flicker-photometry (Lee, Martin, & Valberg, 1988). With this measure, we eliminated any subjective foreground-background contrast differences between the target and the distractors ensuring that participants could not have used this difference to locate the target among the distractors. As a consequence, participants had to rely on the non-singleton color feature to search and locate the targets.

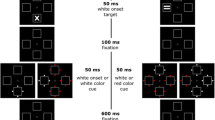
Depicted is a schematic trial in Experiment 1. A black ring was shown as an abrupt onset cue. Sixteen milliseconds after the cue, two additional placeholders appeared. The color target and the distractors appeared after a variable SOA, either along with the two additional placeholders (short SOA) or 1 s after the placeholders (long SOA). The arrow symbolizes the direction of the flow of time. Stimuli are not drawn to scale
Our expectations were straightforward. If in these conditions a validity effect would be found, we can conclude that subliminal cues can capture attention in a truly stimulus-driven way. In contrast, if in these conditions no cue validity effect emerges then we have to conclude that subtle top-down effects may play a role in capture by subliminal onset cues.
To determine whether inhibition of return (IOR) emerged under these very well-controlled circumstances, we also included a cue-target SOA of 1,016 ms. Note that Mulckhuyse et al. (2007), who were the first to use this type of subliminal cueing procedure, did find IOR; their response times were faster for invalid than valid cue conditions for the long cue target SOAs (Klein, 2000). According to Mulckhuyse and Theeuwes (2010a), finding an IOR effect is a strong marker for the stimulus-driven nature of attention capture. Because several attempts to replicate the IOR effect were unsuccessful (Fuchs & Ansorge, 2012a; Fuchs et al., 2013; see also McCormick, 1997), we tried to look at this effect again but now under very well-controlled conditions. We also included many participants (n = 36) to ensure that previous failed attempts were not due to insufficient power.
After the main experiment in which participants performed the target discrimination task, we ran a cue localization task to assess the participants’ awareness of the cues. The performance in this objective cue-localization task was used to calculate the validity effect as a function of the cue’s visibility. This test regresses the individual validity effects on the individual cue visibility measures. In this regression test, two findings indicate a likely origin of the validity effect in subliminal perception: a significant positive intercept at zero visibility and a nonsignificant slope of the regression (Greenwald, Draine, & Abrams, 1996). The corresponding analyses also were conducted.
Methods
Participants
Thirty-six participants (24 females, mean age: 23.3 years) took part. Participants were mostly students and participated on a voluntary basis in return for course credit. They had normal or corrected-to-normal vision, and normal color-vision as assessed by Ishihara color plates. The procedure was explained prior to data acquisition, and informed consent was obtained from each participant.
Apparatus and stimuli
The stimuli were presented on a 19-inch CRT monitor, with a resolution of 1,024 × 768 pixels and a refresh rate of 120 Hz. Head position and a viewing distance of 64 cm were supported by a chin rest with a forehead strip. Stimulus presentation was controlled using Experiment Builder software (SR Research, Kanata, ON, Canada). We used two methods for equating either the luminance or the lightness of the colors. First, CIE-Lab color coordinates were objectively measured, and all colors were calibrated so that they had the same objective luminance as the background (CIE-Lab color coordinates: 35.3/8.4/-22.4): red (35.3/49.5/40.9), green (35.3/-44.4/32.7), and blue (35.3/63.0/-118.9). Second, a flicker photometry technique was used to ensure subjective equi-lightness. Participants were presented with two alternating fields: one in gray (as the background of the monitor), and one in red, green, or blue. The alternation frequency was set to 16 Hz and could be individually adjusted if necessary. Participants could increase and decrease the lightness of the colors stepwise (always by one lightness, L*, unit in the CIE-Lab color space) until the flicker perception was minimal. This procedure was repeated several times until a reproducible value of subjective equi-lightness was found for each color. Because the participant’s performance in flicker photometry can vary between the eyes due to differences of the receptor mosaic and the distribution of macular pigment (Snodderly et al., 2004), the whole experiment, including the flicker photometry technique, was run with the nondominant eye patched.
Procedure
Figure 1 shows the procedure, which is a variation of the paradigm originally developed by Mulckhuyse et al. (2007) and later adapted by Fuchs and colleagues (Fuchs & Ansorge, 2012a, 2012b; Fuchs et al., 2013). Participants viewed the stimuli with the dominant eye (as assessed by the Miles test). Participants worked through two tasks and four blocks: first, two blocks of the target discrimination task, and, second, two blocks of the cue localization task. The target discrimination task was always administered first and it was concluded before the cue localization task. Between the first and the second block, and once again between the third and the fourth block, it was varied whether the targets were presented in objectively equated luminance or in subjectively equated lightness, with the block order balanced across participants (but being the same for each participant in the target discrimination task and in the cue localization task).
Throughout the experiment, eye movements were controlled with the EyeLink 1000 (SR Research, Mississauga, Ontario, Canada) at 1,000 Hz. The proper signal was controlled prior to each trial and the eye tracker recalibrated if necessary. Participants were asked to keep their fixation at the screen center throughout each trial. If the participant’s eye gaze deviated more than 3° from the screen center, the trial was interrupted and the participants received feedback. We counted those trials as “saccade-trials” and repeated them at a later point of the experiment.
In each block, to initiate a trial, participants had to press the #5-key on the number pad of a standard keyboard. Next, a fixation screen occurred. After fixation, a blank screen was shown for 700 ms. Then the cue was presented for 16 ms to the left or right of fixation before two additional black placeholders of the same size and luminance appeared, one at fixation and the other one on the opposite side (Fig. 1). Participants viewed the lateral stimuli at an eccentricity of 11°. The abrupt-onset cue and the placeholders were black 3.9°-diameter rings of 0.4° width (CIE values: 9.6/7.4/-12.1). In the short SOA condition, the color target and two distractors appeared along with the placeholders, whereas in the long SOA condition, the target and the two distractors were presented with a delay of 1 s after the placeholders. The target and the distractors were 2.7°-diameter Landolt rings of 0.6° width, centered inside the cue and the placeholders. The target and the distractor Landolt rings were oriented towards (i.e., open at) their upper, lower, left, or right side. In each trial, the target and the distractors were presented in different colors and with different orientations. For example, when the target was a red ring with its opening towards the top, the two distractors might have been one green ring opening towards the left, and one blue ring opening towards the bottom (Fig. 1). The target screen with the distractors was presented for 400 ms. Next, a blank screen occurred and participants had another 1 s to respond.
In the target discrimination task, participants searched for one particular target color (target colors were balanced across participants) throughout all trials and responded to its orientation as correctly and quickly as possible. At the start of each trial, participants had their right index finger on the #5-key of the number pad and subsequently indicated the orientation of the target by a key press: an orientation towards the top by pressing the #8-key, to the bottom by the #2-key, to the left by the #4-key, and to the right by the #6-key. If the answer was too slow (>1.4 s) or incorrect, participants received written feedback on the screen, and the trial was repeated at a later point in the experiment. Target orientations (upwards, downwards, leftwards, or rightwards), target positions (left or right), cue validity steps (valid or invalid), and SOAs (short: 16 ms or long: 1016 ms) were all equally probable and pseudo-randomized across trials. Distractors’ orientations (upwards, downwards, leftwards, or rightwards) also were pseudo-randomized, with the limitation that distractor orientations were always different from one another and from the target orientation. RT and accuracy were measured. Before data recording, participants got practice until they reached a criterion of 90 % accuracy (9 of 10 trials correct). The two methods of color luminance/lightness equation were tested in separate blocked conditions of 128 trials. In total, subjects had to perform 256 target discrimination trials.
In the cue localization task, cue awareness was assessed with the same participants in a separate block after the target discrimination task. Participants had to report at which of two positions the cue was presented. They had to press a left-hand button with the left index finger if the cue was on the left and a right-hand button with the right index finger if the cue was on the right. The cue localization task was conducted in four mini-blocks consisting of 24 trials each (96 trials in total), immediately following the target discrimination task.
Results
Target discrimination task
RTs
Trials with incorrect responses and misses (1.77 %) and trials with eye gaze deviations from the screen center (5.48 %) were removed for the analysis of the RTs. For every participant, the overall mean and standard deviation of the correct RTs was computed and outliers (RTs deviating more than two standard deviations from the mean) were excluded (4.3 %).
For each participant, mean RTs were computed separately for different conditions so that a mixed-measures ANOVA, with the repeated measurement variables luminance/lightness control (objectively vs. subjectively equated), cue validity (valid vs. invalid), SOA (16 ms vs. 1,016 ms), and visual hemifield of the cue (nasal vs. temporal)Footnote 1, and the between-participants variable target-color (red vs. green vs. blue) could be conducted. The main effect of cue validity was not significant F(1, 33) = 3.27, p = 0.08, ηp 2 = 0.09, but we found a significant two-way interaction between the variables cue validity and SOA, F(1, 33) = 14.63, p < 0.01, ηp 2 = 0.31 (Fig. 2). Subsequent planned one-sided t-tests revealed that at the short SOA, participants were faster when the cue was valid (RT = 678 ms) than when it was invalid (RT = 686 ms), t(35) = 4.47, p < 0.01. At the long SOA, there was a nonsignificant trend towards the reversed effect: participants were faster when the cue was invalid (RT = 694) than when it was valid (RT = 698 ms), t(35) = 1.49, p = 0.073), signifying a trend for the occurrence of IOR. In addition, there was a significant main effect of SOA, F(1, 33) = 17.27, p < 0.01, ηp 2 = 0.35, indicating that participants were faster at the short SOA (RT = 682 ms) than at the long SOA (RT = 695 ms). The remaining main effects and interactions were not significant (all non-significant Fs < 2.96, all nonsignificant ps > 0.07).

Results of Experiment 1, target discrimination task. A Mean reaction time (RT) in milliseconds to targets at cued locations (valid) and to targets at uncued locations (invalid) at the short and at the long SOA. B Mean of the cueing effects (mean RT of the invalid conditions minus mean RT of the valid conditions) with 90 % confidence interval at the short SOA and at the long SOA
Error rates
We counted trials with incorrect responses (i.e., incorrect reports of target orientation and misses) as error trials. In the case of an error, the trial was repeated at a later point of the experiment. Out of all trials, on average 1.77 % were error-trials. Next, we calculated the ratios of error-trials divided by the total number of trials in each condition and performed arcsine-square root transformations of the ratios. A repeated-measurements ANOVA on the transformed data with variables as above led to no effects that would be indicative of a speed accuracy trade-off in the cueing effects. There was only a significant main effect of the variable SOA, F(1,35) = 4.40, p < 0.05, ηp 2 = 0.11, with on average more errors at the short SOA (2.10 %) than at the long SOA (1.44 %), and a significant main effect of the variable visual hemifield, F(1,35) = 4.13, p < 0.05, ηp 2 = 0.11. Other effects were not significant (all nonsignificant Fs < 2.92, all nonsignificant ps > 0.10).
Cue localization and the regression of validity effects on cue visibility
After the target discrimination task, participants were informed that in each trial a cue had appeared 16 ms earlier than the two other placeholders. Participants were asked whether they had seen this cue and reported unawareness of the cue in the target discrimination task, indicating subjective unawareness of the cues, and hence subliminal processing (Merikle, Smilek & Eastwood, 2001). Objective cue visibility was assessed with the same participants based on their performance in the cue localization task. For our analysis, we regarded correct reports as hits and incorrect reports as false alarms (FAs), and d’ was calculated as the difference between the z-transformed probabilities of the hits minus the z-transformed probabilities of the FAs. The resulting visibility index d’ should be close to zero in case participants are performing truly at chance probability.
Mean d’ was 0.34 and, though low, significantly different from zero or chance-level performance, t(35) = 4.98, p < 0.01. No bias towards one or the other judgment (left or right) was observed. To assess the cue’s validity effect at the short SOA under zero discrimination conditions, we calculated regressions of the individual validity effects (calculated as invalid RTs minus valid RTs in the short-SOA conditions) on the individual cue localization indices (d’) of the short SOA conditions (Fig. 3). A significant intercept of the regression at zero cue visibility indicated that the RT cueing effect was significantly above zero (a = 7.30, p < 0.05) when d’ was equal to zero. We found no significant correlation between discrimination performance and cueing effect (beta = 4.60, p = 0.21).

Experiment 1: Scatter plot showing for each participant the individual RT cueing effects (invalid RT minus valid RT) on the y axis as a function of an individual’s visibility index (on the x axis). The panel shows the results in the short SOA conditions. The solid line shows the regression line of the cueing effect on discrimination (d’), with its 95 % confidence interval in red. The intercept at the point of zero discrimination is significantly different from zero (a = 7.30; p < 0.05)
Discussion
Experiment 1 suggests that subliminal onset cues capture attention in a stimulus-driven way. In particular, we ruled out influences of two kinds of possible top-down attentional control settings: 1) By presenting non-singleton targets, a search setting for singletons (Bacon & Egeth, 1994) was prevented, and 2) by equating the target and distractor’s objective luminance and subjective lightness to the background, a search setting for foreground/background contrast was prevented. Yet, even though we ensured that the most subtle top-down effect could not play a role, we still found a robust validity effect, with faster responses in the valid than in invalid cue conditions in the short-SOA condition.
Consistent with the claim of stimulus-driven capture is the finding that the cueing effect reversed at longer SOAs. Indeed, comparable to the effect size reported by Mulckhuyse and Theeuwes (2010a) there was a small (4 ms) IOR effect which in the current study was trending towards significance. As argued, the occurrence of IOR is a true marker of stimulus-driven attentional capture (Mulckhuyse et al., 2007; Schreij, Theeuwes, & Olivers, 2010) and the current data pattern is consistent with this view. As different tasks were used in these experiments, it may not be surprising that in the current experiment there was only a trend towards IOR while in Mulckhuyse et al. (2007) IOR was reliable. The current experiment used a discrimination task while Mulckhuyse et al. (2007) used a detection task. It is well known that IOR is less robust with discrimination responses compared with detection responses (Egly, Rafal, & Henik, 1992; Terry, Valdes, & Neill, 1994). It also should be noted that some previous studies also have failed to find IOR with subliminal cues (Fuchs & Ansorge, 2012a , Fuchs et al., 2013; McCormick, 1997), which may question the claim that IOR is the hallmark of stimulus-driven capture (Fuchs & Ansorge, 2012a). Indeed, some studies have found IOR in a task that involved contingent top-down set (e.g., Ansorge, Priess, & Kerzel, 2013), whereas others failed to find IOR in conditions of contingent capture (Schreij et al., 2010).
Like in previous experiments (e.g., Fuchs & Ansorge 2012a, b), cue discrimination was too high to pass a criterion of objective subliminality. Yet, all participants reported having been subjectively completely unaware of the cue after the target discrimination task indicating subjective subliminality of the cues (Merikle et al., 2001). As an additional marker of subliminal processing, we found a significant intercept at zero visibility in a regression of the validity effect on cue visibility that indicated capture of attention even for clearly subliminal cues (Greenwald et al., 1996). In addition, the correlation between the validity effect and cue visibility was not reliable which suggests that the cue visibility is not responsible for the cueing effect observed. Together, these results suggest that the validity effect we observed was likely due to subliminal attentional capture by the onset cue.
Experiment 2
Although our participants in Experiment 1 regularly reported that they did not see the cues during the target discrimination task, they performed better than chance on the objective cue localization task. Certainly, it is possible that to some extent participants could “see” the cue and simply applied a too conservative criterion when judging their awareness. However, to achieve a very sensitive measure of residual cue visibility we might have inadvertently created a visibility test which was also sensitive for the awareness-independent processing of the cues (Reingold & Merikle, 1988). In particular, the cue localization task of Experiment 1 fulfilled the criteria for an awareness-independent motor activation effect by the cues. That is, the required response was always a left-hand key for left cues and aright-hand key for right cues. With this kind of fixed stimulus to response mapping, it is very well possible that the cues primed the corresponding motor response even when they actually remained below the threshold of awareness (Klotz & Neumann, 1999). In addition, during the experiment cue visibility would have suffered from the requirement to search and discriminate the targets. This, however, was not the case in the cue visibility test in which the participants could fully concentrate on the cues. So, it is possible that the cue visibility test in Experiment 1 overestimated cue visibility during target localization.
To test these notions, we assessed cue visibility in Experiment 2 in a combined target localization/cue discrimination task to find out how well participants are able to “see” the cue when they are concurrently engaged in the task searching for and discriminating a target. Crucially, participants did not report cue location by pressing a left-hand button for left cues and a right-hand button for right cues as in Experiment 1. In Experiment 2, after a target response participants were randomly presented with one cue-to-response mapping rule that required pressing either the upper key for a right cue and the lower key for a left cue, or the upper key for a left and the lower key for a right cue. With this kind of random stimulus to response mapping, awareness-independent activation of the required motor response by the cue can be definitely ruled out (Neumann & Klotz, 1994). Finally, Mulckhuyse et al. (2007) had reported chance performance in their cue visibility test with a different luminance of the cues than we had used in our Experiment 1. Therefore, in Experiment 2 the luminance of our stimuli was adjusted to be basically the same as in Mulckhuyse et al.
Like in Experiment 1, the target discrimination task was run and was completed before cue visibility was assessed. We introduced a neutral cue-absent condition to learn more about the origin of the validity effect. So far, the validity effect of subliminal onset cues has been measured as an RT advantage in the valid condition relative to the invalid condition, with cue and target on opposite sides. Basically, the validity effect could thus reflect facilitation at the cued location, impediments by a cue at the opposite location of the target, or both of these effects. With the changed protocol, we should be able to 1) replicate the validity effect at the short SOA of Experiment 1, and 2) assess attentional capture relative to a neutral condition in which no cue was presented. If capture of attention by the cue facilitates searching for the target, we expected to find an advantage in cued relative to neutral conditions. Moreover, this manipulation allowed us to compare a neutral condition with invalid cue conditions. This may turn out to be important as RT costs (RT disadvantage in invalid cue conditions relative to neutral conditions) are only observed if attention is “actively” shifted to the cue in a top-down manner. If the capture of attention is truly automatic and stimulus driven we expect only RT benefits and no RT costs (Jonides & Mack, 1984; Posner & Snyder, 1975).
Methods
Participants
Twelve new participants (9 females, mean age: 22.42 years) took part in Experiment 2.
Apparatus and stimuli
In order to approximate the conditions of Mulckhuyse et al. (2007), the target, the distractors, and the screen background were of 5.2 cd/m2 luminance. The screen background (CIE-Lab: 27.6/10.3/-21.2) and the objectively equiluminant color stimuli red (CIE-Lab: 27.4/38.9/25.4), green (CIE-Lab: 27.3/-33.6/21.8), and blue (CIE-Lab: 27.5/49.9/-96.4) were darker than in Experiment 1. Like in Experiment 1, flicker photometry was used to ensure subjective equi-lightness. Again, the whole experiment, including flicker photometry, was run with the nondominant eye patched.
Procedure
Participants performed two tasks: the first two blocks consisted of the target discrimination task, and the second two blocks consisted of the combined target discrimination/cue localization task (which was different from the one used in Experiment 1). Between the first and the second block, and once again between the third and the fourth block, the targets were either presented in objectively equated luminance or in subjectively equated lightness (balanced across participants).
The procedure in the two blocks of the target discrimination task was the same as in Experiment 1, with the exceptions that 1) a cue was presented only in 66 % of the trials (33 % were neutral trials in which the cue was absent), and 2) we no longer employed a long SOA. Also different from Experiment 1, the cue and the placeholders were brighter than the background. The background had a luminance of 5.2 cd /m2, the cue and the placeholders had a luminance of 13 cd/m2 (CIE-Lab: 42.7/10.4/-27.3). Again, the two methods of equating color luminance were tested in separate blocks of 132 trials. In total, subjects had to correctly perform on 264 trials of the target discrimination task. If an error was made, the trial was repeated at a later point of the experiment.
In the combined target discrimination/cue localization task, the target discrimination part was the same as in the preceding blocks. Participants had to correctly discriminate a target on a total of 240 trials, and incorrect trials were repeated at a later point of a block. On altogether 60 trials, if (1) a cue had been presented and (2) the response to the target was correct, the participants were asked subsequently whether the cue had appeared on the left or on the right. On these cue localization trials, participants had to report cue location by pressing the #2-key and the #8-key on the number pad with their right index finger. “Left cue” and “right cue” were randomly assigned to the two keys on each cue-localization trial. To that end, immediately after participants had reported target identity in a cue-localization trial, a screen appeared indicating the corresponding key assignment. There was no time constraint for reporting cue location (Fig. 4). Altogether, there were two blocks of cue discrimination, one with objectively and one with subjectively equated luminance/lightness.

Depicted is a schematic trial in Experiment 2. A white ring was shown as an abrupt onset cue in 66 % of the trials; 33 % of the trials did not contain a cue. Sixteen milliseconds after the cue, the target screen appeared with all three placeholders, the target and two distractors. In the combined target discrimination/cue localization task, cue visibility was assessed in 25 % of the trials immediately after the target discrimination had been made. The arrow symbolizes the direction of the flow of time. Stimuli are not drawn to scale
Results
Target discrimination task
RTs
Trials with incorrect responses (3.66 %) and trials with saccades (4.82 %) were excluded from the analysis of the RTs. For each participant, the overall mean and standard deviation of the correct RTs was computed and outliers (RTs deviating more than two standard deviations from the mean) also were excluded (4.4 %).
Next, mean RTs were computed separately for different conditions so that a repeated-measures ANOVA, with the variables luminance/lightness control (objective vs. subjective) and cue validity (valid vs. invalid vs. neutral) could be computed. It revealed a highly significant main effect of cue validity, F(2, 22) = 16.40, p < 0.01, ηp 2 = 0.60. RTs in the valid condition were significantly faster (669 ms) than in the invalid [688 ms, t(11) = 4.3, p < 0.01] and the neutral condition [687 ms, t(11) = 5.3, p < 0.01], whereas the RT difference between the invalid condition and the neutral condition was not significant [t(11) = 0.37, p = 0.36]. All other effects of the ANOVA were not significant (all nonsignificant Fs < 2.19, all nonsignificant ps > 0.14). The results are depicted in Fig. 5.

Results of Experiment 2, target discrimination task. A Mean RT in milliseconds to targets at cued locations (valid), to targets at uncued locations (invalid) and to targets in cue-absent trials (neutral). B Means of the overall validity (cueing) effect (RTs in invalid minus valid conditions), the benefit (RTs in neutral minus valid conditions), and the cost (RTs in the neutral minus invalid conditions) with the 90 % confidence interval
Error rates
Of all trials, 3.66 % were error trials. Like in Experiment 1, we calculated the ratios of error trials in each condition and performed an arcsine-square root transformation. A repeated-measurements ANOVA with the variables luminance/lightness control (subjective vs. objective) and cue validity (valid vs. invalid vs. neutral) revealed no significant effects (all nonsignificant Fs < 1.96, all nonsignificant ps > 0.17).
Cue localization and the regression of validity effects on cue visibility
After the first two blocks of the target discrimination task, participants were informed about the cue. Again participants reported subjective unawareness of the cue in the preceding target discrimination task. To assess cue visibility objectively, we conducted a combined target discrimination/cue localization task. We regarded correct reports of the cue’s location as hits and incorrect reports of the location as false alarms (FAs), and d’ was calculated as the z-transformed probabilities of the hits minus the z-transformed probabilities of the FAs. Mean d’ was −0.06 and not significantly different from zero, t(11) = −0.60, p = 0.56. No bias towards one or the other judgment (left or right) was observed. To assess the cue’s validity effect under zero discrimination conditions, we again calculated regressions of the individual validity effects (calculated as invalid RTs minus valid RTs) on the individual cue localization indices (d’). The intercept of the regression at zero cue visibility was significantly different from zero (a = 18.44, p < 0.01), and no significant correlation between discrimination performance and cueing effects was found (beta = −2.59, p = 0.87). Figure 6 shows a scatter plot of the data and the regression.

Experiment 2: Scatter plot showing for each participant the individual RT cueing effects (invalid RT minus valid RT) on the y axis as a function of an individual’s visibility index (on the x axis). The solid line shows the regression of the cueing effect on discrimination (d’), with its 95 % confidence interval in red. The intercept is significantly different from zero (a = 18.44; p < 0.01)
Discussion
Experiment 2 replicated the cue validity effect as RTs were faster in the valid than in the invalid cue condition. Because Experiment 2 also had a neutral condition, we were able to examine RT costs and benefits. As shown in Fig. 5, there were only RT benefits and no costs: relative to the neutral condition there was a cueing effect for valid cue conditions, and no cueing effect for the invalid cue conditions. As previous findings have shown that this pattern of results (RT benefits and no costs) only occurs when attention is controlled in a truly automatic and stimulus-driven way (Jonides & Mack, 1984; Posner & Snyder, 1975), the present findings further support our conclusion that the capture of attention by the subliminal cue was stimulus-driven without the involvement of any top-down set.
Regarding cue visibility, Experiment 2 supports the assumption that the cue remained below the threshold of the participants’ awareness when it captured attention. Again, participants indicated that they were completely unaware of the cues. Different from Experiment 1, also the objective visibility test indicated no awareness of the cue. When we assessed objective cue visibility in a combined target discrimination/cue localization task, performance on cue localization was at chance level. This also is in line with the results of the regression of the visibility index d’ on the cueing effect. Like in Experiment 1, we found a significant intercept at the point of zero discrimination and no correlation between the cueing effect and the cue visibility.
Although a single cue localization task may have been the more conservative method, we think that the visibility assessment in Experiment 2 is more appropriate, because it better reflects whether participants became aware of the cue when they were engaged in the target discrimination task. Furthermore, in Experiment 2, we ensured that possible awareness-independent motor priming by the cues could not play a role. Together the results of Experiment 2 show that spatial attention was shifted to the abrupt onset cue in a stimulus-driven way. Participants, however, remained unaware of the cue and could not indicate which location contained the abrupt onset.
General Discussion
We tested whether subliminal onset cues capture attention in a stimulus-driven way or whether capture is contingent on top-down attentional control settings. The current findings unequivocally provide evidence that subliminal attentional capture is completely stimulus-driven. Answering this question is important as previous studies provided evidence that the validity effect of subliminal onset cues depended on the top-down control settings (e.g., Fuchs & Ansorge, 2012b; Scharlau & Ansorge, 2003), whereas others claimed that subliminal abrupt onsets capture attention in a pure stimulus-driven way (McCormick, 1997; Mulckhuse et al., 2007). In particular, past research allowed two types of top-down contingent capture to have a possible effect: (1) participants were searching for a target singleton while the abrupt onset cue also was a singleton (singleton vs. feature search mode cf. Bacon & Egeth, 1994), and (2) participants may have searched for a target having particular contrast while the cue also may have had this contrast. The current study ruled out both of these possible top-down attentional control settings by presenting the target as a nonsingleton and by equating the objective luminance and subjective lightness between targets, distractors, and the background. As is clear from the data, we found a cue validity effect with faster responses in cue valid conditions relative to cue invalid conditions (Experiments 1 and 2) and also faster responses in cue valid conditions relative to the cue-absent neutral conditions (Experiment 2) at the short-SOA. This finding clearly supports the conclusion that the onset cues captured attention in a stimulus-driven way. The results of Experiment 2 in which we observed RT benefits and no costs further support the conclusion that capture of attention by subliminal cues is truly automatic and stimulus-driven. The trend in finding IOR in Experiment 1 also is consistent with this claim, as some have claimed that IOR is the hallmark of exogenous stimulus-driven capture (Mulckhuse et al., 2007; Theeuwes & Chen, 2005).
Cue visibility
Like in previous experiments (e.g., Fuchs & Ansorge 2012a, b), cue discrimination was too high to pass a criterion of true objective subliminality in Experiment 1. Nonetheless, we think that the cueing effect is subliminal in origin. One argument for this is the result of the regression of the validity effect on cue visibility. In both experiments, we found a significant intercept at zero visibility in a regression of the validity effect on cue visibility that indicated capture of attention even for subliminal cues (Greenwald et al., 1996). The same analyses showed that there was no significant correlation between the cueing effect and cue visibility as would have been expected if cue visibility were responsible for the capture of attention.
Furthermore, cue visibility was maybe overestimated in Experiment 1 because participants could fully focus on cue discrimination during the cue localization task, whereas during the experiment cue visibility could have suffered from the requirement to search for and discriminate the targets at the same time. In addition, the visibility task of Experiment 1 fulfilled the criteria for an awareness-independent motor activation effect by the cues (Klotz & Neumann, 1999), because there was a fixed stimulus to response mapping. That is, reports for right cues were always given by a right-hand key press and for left cues by a left-hand key press. In Experiment 2, we eliminated these concerns. First, we varied the required response for cues on either side from trial to trial so that the stimulus to response mapping was random. Second, we assessed cue visibility in a combined target discrimination/cue localization task to test what participants are actually able to “see” when they are concurrently engaged in the target discrimination task. Mean cue discrimination under these conditions was now at chance level. Together, the results suggested that the validity effect we observed in both experiments was most likely due to subliminal capture by the cues. This implies that stimulus-driven shifts of spatial attention are indeed possible even without the awareness of which location is attended.
Stimulus-driven capture under subliminal and top-down contingent capture under supraliminal conditions?
It also is important to consider the reason for (1) the absence of top-down contingent capture by abrupt onsets with the presently used cues but (2) the presence of top-down contingent capture under supraliminal cueing conditions (Folk et al., 1992). Although it is unclear whether attention capture by supraliminal abrupt onsets is entirely and exclusively driven by top-down influences (e. g., Belopolsky, Schreij, & Theeuwes, 2010; Liao & Yeh, 2013; Schreij, Owens & Theeuwes, 2008), some of our recent results suggest that the contribution of top-down contingent capture to a net cueing effect could at least be much stronger in supraliminal conditions than in subliminal conditions. For example, in Fuchs et al. (2013), supraliminal and weak (subliminal) abrupt onset cues were compared by their ability to capture attention. It was found that (1) subliminal abrupt onsets led to more capture than supraliminal onsets and (2) that only with the supraliminal onsets, capture was much stronger (and exclusively observed) for onset cues that matched the top-down attentional control settings. In contrast, the supraliminal cues that did not match the top-down control settings produced almost no validity effect.
Jointly, these findings suggest that the contingent cueing that requires supraliminal conditions involves different attentional processes than those involved in subliminal cueing. As has been argued before (Theeuwes, 2010), attentional capture that is contingent on what people are looking for, should not be labeled as exogenous. Indeed, when attention moves to the cue that matches the feature observers are looking for (as is typically the case in contingent capture studies), this shift of attention is completely top-down. If features do not match the top-down set, it may not be surprising that there is no capture for a mismatching feature as this does not fit the top-down set. In case of subliminal cues, our research shows that top-down sets cannot be as efficiently used to shift attention as the features that are supposed to drive the capture are not perceived by the participant. When the cue is weak or subliminal, it is simply not registered so that the participants remain unaware of the fact that these cues lead them astray in invalid conditions. As a consequence, a weak onset cue is not suppressed and leads to a stronger validity effect that also does not depend on the exact features of this cue. On this view, there might be a stimulus-driven mechanism to attend to abrupt onsets in the periphery. However, this mechanism can be overruled by active suppression once cues are clearly noted, so that this mechanism does no longer ensure the capture of attention by abrupt onsets in each and every condition. Nonetheless, evidence for an evolutionary default mode of stimulus-driven capture by abrupt onsets can still be found with weak cues or in subliminal cueing conditions.
Notes
The variable hemifield was included because with eye patching the cues (and targets) were presented to the nasal or to the temporal visual hemifield alone. Assuming that the cueing effect or IOR originates at collicular levels, more cueing or IOR might be found with temporally than with nasally presented cues (Ansorge, 2003; Mulckhuyse & Theeuwes, 2010b; but see Fuchs & Ansorge, 2012a) because of the stronger crossed than the weaker uncrossed pathway of the retino-collicular projection of the visual system (Rafal, Henik, & Smith, 1991). However, in the current study, this variable had no effect on subliminal cueing.
References
Ansorge, U. (2003). Asymmetric influences of temporally vs. nasally presented masked visual information: Evidence for collicular contributions to nonconscious priming effects. Brain and Cognition, 51(3), 317–325. doi:10.1016/S0278-2626(03)00034-4
Ansorge, U., Priess, H.-W., & Kerzel, D. (2013). Effects of relevant and irrelevant color singletons on inhibition of return and attentional capture. Attention, Perception, & Psychophysics, 75(8), 1687–1702. doi:10.3758/s13414-013-0521-2
Bacon, W. F., & Egeth, H. E. (1994). Overriding stimulus-driven attentional capture. Perception & Psychophysics, 55(5), 485–496. doi:10.3758/BF03205306
Belopolsky, A. V., Schreij, D., & Theeuwes, J. (2010). What is top-down about contingent capture? Attention, Perception, & Psychophysics, 72(2), 326–341. doi:10.3758/APP.72.2.326
Breitmeyer, B. G., & Ganz, L. (1976). Implications of sustained and transient channels of visual pattern masking, saccadic suppression, and information processing. Psychological Review, 83(1), 1–16. doi:10.1037/0033-295X.83.1.1
Egly, R., Rafal, R. D., & Henik, A. (1992). Reflexive and voluntary orienting in detection and discrimination tasks. St. Louis: Poster presented at the meeting of the Psychonomic Society.
Folk, C. L., Remington, R. W., & Johnston, J. C. (1992). Involuntary covert orienting is contingent on attentional control settings. Journal of Experimental Psychology: Human Perception and Performance, 18(4), 1030–1044. doi:10.1037/0096-1523.18.4.1030
Fuchs, I., & Ansorge, U. (2012a). Inhibition of return is no hallmark of exogenous capture by unconscious cues. Frontiers in Human Neuroscience, 6(30), 1–8. doi:10.3389/fnhum.2012.00030
Fuchs, I., & Ansorge, U. (2012b). Unconscious cueing via the superior colliculi: Evidence from searching for onset and color targets. Brain Sciences, 2(1), 33–60. doi:10.3390/brainsci2010033
Fuchs, I., Theeuwes, J., & Ansorge, U. (2013). Exogenous attentional capture by subliminal abrupt-onset cues: Evidence from contrast-polarity independent cueing effects. Journal of Experimental Psychology: Human Perception and Performance, 39(4), 974–988. doi:10.1037/a0030419
Greenwald, A. G., Draine, S. C., & Abrams, R. L. (1996). Three cognitive markers of unconscious semantic activation. Science, 273, 1699–1702. doi:10.1126/science.273.5282.1699
Gunther, K. L., & Dobkins, K. R. (2002). Individual differences in chromatic (red/green) contrast sensitivity are constrained by the relative number of L- versus M-cones in the eye. Vision Research, 42(11), 1367–1378. doi:10.1016/S0042-6989(02)00043-3
Held, B., Ansorge, U., & Müller, H. (2010). Masked singleton effects. Attention, Perception & Psychophysics, 72(8), 2069–2086. doi:10.3758/APP.72.8.2069
Jonides, J. (1981). Voluntary versus automatic control over the mind’s eyes movement. In J. B. Long & A. D. Baddeley (Eds.), Attention and Performance IX (pp. 187–203). Hillsdale: Erlbaum.
Jonides, J., & Mack, R. (1984). On the cost and benefit of cost and benefit. Psychological Bulletin, 96(1), 29–44. doi:10.1037//0033-2909.96.1.29
Klein, R. M. (2000). Inhibition of return. Trends in Cognitive Sciences, 4(4), 138–147. doi:10.1016/S1364-6613(00)01452-2
Klotz, W., & Neumann, O. (1999). Motor activation without conscious discrimination in metacontrast masking. Journal of Experimental Psychology: Human Perception and Performance, 25(4), 976–992. doi:10.1037/0096-1523.25.4.976
Leber, A. B., & Egeth, H. E. (2006). It’s under control: Top-down search strategies can override attentional capture. Psychonomic Bulletin & Review, 13(1), 132–138. doi:10.3758/BF03193824
Lee, B. B., Martin, P. R., & Valberg, A. (1988). The physiological basis of heterochromatic flicker photometry demonstrated in the ganglion cells of the macaque retina. Journal of Physiology, 404, 323–347.
Liao, H.-I., & Yeh, S.-L. (2013). Capturing attention is not that simple: Different mechanisms for stimulus-driven and contingent capture. Attention, Perception, & Psychophysics, 75(8), 1703–1714. doi:10.3758/s13414-013-0537-7
McCormick, P. A. (1997). Orienting attention without awareness. Journal of Experimental Psychology: Human Perception and Performance, 23(1), 168–180. doi:10.1037//0096-1523.23.1.168
Merikle, P. M., Smilek, D., & Eastwood, J. D. (2001). Perception without awareness: Perspectives from cognitive psychology. Cognition, 79(1–2), 115–134. doi:10.1016/S0010-0277(00)00126-8
Mulckhuyse, M., & Theeuwes, J. (2010a). Unconscious attentional orienting to exogenous cues: A review of the literature. Acta Psychologica, 134(3), 299–309. doi:10.1016/j.actpsy.2010.03.002
Mulckhuyse, M., & Theeuwes, J. (2010b). Unconscious cueing effects in saccadic eye movements – facilitation and inhibition in temporal and nasal hemifield. Vision Research, 50(6), 606–613. doi:10.1016/j.visres.2010.01.005
Mulckhuyse, M., Talsma, D., & Theeuwes, J. (2007). Grabbing attention without knowing: Automatic capture of attention by subliminal spatial cues. Visual Cognition, 15(7), 779–788. doi:10.1080/13506280701307001
Neumann, O., & Klotz, W. (1994). Motor responses to nonreportable, masked stimuli: Where is the limit of direct parameter specification? In C. Umiltà & M. Moscovitch (Eds.), Attention and performance XV: Conscious and nonconscious information processing (pp. 123–150). Cambridge: MIT Press.
Posner, M. I. (1980). Orienting of attention. Quarterly Journal of Experimental Psychology, 32A(1), 3–25. doi:10.1080/00335558008248231
Posner, M. I., & Cohen, Y. (1984). Components of visual orienting. In H. Bouma & D. Bouwhuis (Eds.), Attention and Performance X: Control of language processes (pp. 531–556). Hillsdale: Lawrence Erlbaum.
Posner, M.I., & Snyder, C. (1975). Attention and cognitive control. In R. L. Solso (Ed.), Information processing and cognition: The Loyola symposium (pp. 55-86).
Rafal, R., Henik, A., & Smith, J. (1991). Extrageniculate contributions to reflex visual orienting in normal humans: A temporal hemifield advantage. Journal of Cognitive Neuroscience, 3(4), 322–328. doi:10.1162/jocn.1991.3.4.322
Reingold, E. M., & Merikle, P. M. (1988). Using direct and indirect measures to study perception without awareness. Perception & Psychophysics, 44(6), 563–575. doi:10.3758/BF03207490
Scharlau, I., & Ansorge, U. (2003). Direct parameter specification of an attention shift: Evidence from perceptual latency priming. Vision Research, 43(12), 1351–1363. doi:10.1016/S0042-6989(03)00141-X
Schreij, D., Owens, C., & Theeuwes, J. (2008). Abrupt onsets capture attention independent of top-down control settings. Perception & Psychophysics, 70(2), 208–218. doi:10.3758/PP.70.2.208
Schreij, D., Theeuwes, J., & Olivers, C. N. L. (2010). Abrupt onsets capture attention independent of top-down control settings II: Additivity is no evidence for filtering. Attention, Perception, & Psychophysics, 72(3), 672–682. doi:10.3758/APP.72.3.672
Snodderly, D. M., Mares, J. A., Wooten, B. R., Oxton, L., Gruber, M., & Ficek, T. (2004). Macular pigment measurement by heterochromatic flicker photometry in elderly subjects: The carotenoids and age related eye study. Investigative Ophthalmology & Visual Science, 45(2), 531–538.
Terry, K. M., Valdes, L. A., & Neill, W. T. (1994). Does “inhibition of return” occur in discrimination tasks? Perception & Psychophysics, 55(3), 279–286.
Theeuwes, J. (2010). Top–down and bottom–up control of visual selection. Acta Psychologica, 135(2), 77–99. doi:10.1016/j.actpsy.2010.02.006
Theeuwes, J., & Chen, C. Y. D. (2005). Attentional capture and inhibition (of return): The effect on perceptual sensitivity. Perception & Psychophysics, 67(8), 1305–1312. doi:10.3758/BF03193636
Yantis, S., & Jonides, J. (1984). Abrupt visual onsets and selective attention: Evidence from visual search. Journal of Experimental Psychology: Human Perception and Performance, 10(5), 601–620. doi:10.1037//0096-1523.10.5.601
Author Note
Tobias Schoeberl, Faculty of Psychology, University of Vienna, Vienna, Austria.
Correspondence concerning this article should be addressed to Tobias Schoeberl, Faculty of Psychology, University of Vienna, Liebiggasse 5, A-1010 Vienna, Austria. E-mail: tobiasschoeberl11@gmail.com.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Schoeberl, T., Fuchs, I., Theeuwes, J. et al. Stimulus-driven attentional capture by subliminal onset cues. Atten Percept Psychophys 77, 737–748 (2015). https://doi.org/10.3758/s13414-014-0802-4
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13414-014-0802-4