
Abstract
Attentional control over prepotent responses has previously been shown by manipulating the probability with which stimuli appear. Here, we examined whether prepotent responses to self-associated stimuli can be modulated by their frequency of occurrence. Participants were instructed to associate geometric shapes with the self, their mother, or a stranger before having to judge whether the sequential shape–label pairs matched or mismatched the instruction. The probability of the different shape–label pairs was varied. There was a robust advantage to self-related stimuli in all cases. Reducing the proportion of matched self pairs did not weaken performance with self-related stimuli, whereas reducing the frequency of either matched mother or stranger pairs hurt performance, relative to when the different match trials were equiprobable. In addition, while mother and stranger pairs jointly benefitted when they both occurred frequently, there were benefits only to self pairs when the frequency of self trials increased along with either mother or stranger trials. The results suggest that biases favoring self-related stimuli occur automatically, even when self-related stimuli have a low probability of occurrence, and that expectations to frequent, self-related stimuli operate in a relatively exclusive manner, minimizing biases to high-probability stimuli related to other people. In contrast, biases to high-familiarity stimuli (mother pairs) can be reduced when the items occur infrequently and they do not dominate expectations over other high-frequency stimuli.
Similar content being viewed by others

Introduction
Automaticity and prepotent responses
Since the first demonstrations of the Stroop effect (Stroop, 1935), it has been known that stimuli vary in the degree to which they generate prepotent responses. The Stroop effect itself provides one example of a learned prepotent response based on word reading, which operates more quickly and can consequently compete with the response to naming of the word’s color (for a review, see MacLeod, 1991). There also exist other forms of prepotent responses that may reflect more in-built tendencies—for example, to respond to the location where a stimulus originates (e.g., as in the Simon effect; Simon, 1969) or the concordance between the visual properties of objects and the way they are gripped (for evidence of an irrelevant grip modulating responses to both objects and nonobjects, see, e.g., Kumar, Yoon, & Humphreys, 2012; Yoon & Humphreys, 2005). One important issue is whether the influence of these prepotent responses can be controlled.
The ability to control prepotent responses has been examined by manipulating the probability with which differences classes of stimuli appear (e.g., Milliken & Lupiáñez, 2005; Schmidt, 2013). For example, studies of both Stroop and Simon effects show that the degree of interference observed when the response dimensions of the stimuli are incongruent, relative to when they are congruent, varies with the proportion of congruent trials (for work on Stroop interference, see, e.g., Logan, Zbrodoff, & Williamson, 1984; Schmidt & Besner, 2008; for work on the Simon effect, see Stürmer, Leuthold, Soetens, Schröter, & Sommer, 2002). The changes in the magnitude of the congruency effects have received several interpretations. One suggestion is that expectancies are formed on the basis of the prior trial (e.g., whether the stimulus and the response were congruent [Gratton, Coles, & Donchin, 1992] or, even more, the time taken for encoding [Schmidt & De Houwer, 2011]); another is that there is implicit learning of stimulus–response relations that modulates performance (Blais, Harris, Guerrero, & Bunge, 2012; Schmidt, De Houwer, & Besner, 2010). These results suggest that there is at least some adaptation to apparently automatic processes and even that there can be attentional control over the effects of prepotent responses.
Personal significance and prepotent responses
Recent research has demonstrated that self-related stimuli also elicit strong prepotent responses. For example, studies of face processing show that attention can be attracted to participants’ own faces when presented as background distractors (Brédart, Delchambre, & Laureys, 2006; Sui, Chechlacz & Humphreys 2012), disrupting primary task performance. The bias toward self-related stimuli has also been shown using simple associative matching in which a neutral stimulus is linked to a label related to the self versus other people. Sui, He, and Humphreys (2012) assigned three geometric shapes to three people (e.g., self, friend, and stranger) and then had participants judge whether shape–label pairs were correctly or incorrectly matched. Participants were faster and more accurate to match self-related shape–label, as compared with shape–label matches for other people. Furthermore, shape–label matches to self-related stimuli were relatively unaffected by reductions in contrast, whereas effects of contrast reduction were evident for stimuli related to other people, indicating enhanced perceptual processing for self-related items. Unlike the studies of face processing, the self-bias effects were observed when self-related information was linked to neutral shapes and so cannot be attributed to differences in the familiarity of the shapes.
The present study
In the present study, we set out to assess whether there was modulation of the self-bias effect when the proportions of different types of trial were manipulated. Are self-bias effects in perceptual matching modulated by expectancy and/or intertrial learning of stimulus contingencies in the same way as matching performance with stimuli related to other people? To test this, we report four experiments in which we varied the proportions of self- to other-related trials and tested whether the self-bias could be overcome (or even reversed) when self-related stimuli appeared on a minority of trials, relative to other-related stimuli. We used three labels corresponding to the participant (you), to the participant’s mother, or to a stranger. The experiments were conducted with Chinese participants, for whom the mother–self relationship is known to be close (Wang et al., 2012; Zhu & Zhang, 2002; Zhu, Zhang, Fan, & Han, 2007), thus providing a strong test of whether self-related is “special” over and above the effects of the familiarity of the person linked to the label. In addition, we used a sequential version of the shape–label matching task, in which participants saw and made no response to the shape, which was followed by a matching or mismatching label to which they responded. This sequential version ensures that participants can maximize the probability relations in the experiment to predict the response from the initial shape (cf. Schmidt & Besner, 2008). In Experiment 1, there was an equal number of matching trials for the three types of association (you, mother, and stranger), providing baseline data under sequential matching conditions (cf. Sui, He, & Humphreys, 2012; Sui, Rotshtein, & Humphreys, 2013). In Experiment 2, the frequency of occurrence for matched self pairs was reduced, while the frequencies of matched mother and stranger pairs were both increased. We assessed whether there was a decrease in the self advantage here, relative to the baseline condition (in Experiment 1). In Experiment 3, there were decreases in the proportion of matching trials for mother pairs and increases in the probabilities of matching trials for self and stranger pairs. In Experiment 4, we reduced the probability of matching trials for stranger pairs and increased the proportions of matching trials for self and mother pairs. In all the experiments, the proportions of the different shape–label pairings on mismatch trials was held constant; this meant that, in the low-probability match conditions, there was a greater likelihood of a mismatch than of a match trial following the critical shape. We report that, relative to the baseline equal probability condition, decreases in the proportion of self pairs had no effect on the self advantage, although the bias was enhanced as the proportions of self-matching trials increased. In contrast, having mother pairs appear on a minority of trials eliminated the normal advantage for mother over stranger pairs (see Sui, He, & Humphreys, 2012). The contrasting effects for self and for mother pairs provide important evidence that there are least nonlinear effects of familiarity or that self-biases are not due to familiarity at all. In addition, while increasing the probability of both mother- and stranger-related trials facilitated performance in both conditions, increasing the proportion of self-related trials had an exclusive benefit, minimizing the gains from increasing the probability of other trial types. Thus, self-related stimuli not only produce automatic benefits on performance (under low-probability conditions), but also dominate performance when their probability of occurrence increases. We discuss the implications for understanding self-bias and cognitive control.
Method
Participants
Twenty-two Chinese college students from Tsinghua University (7 male; age range, 20–27 years; mean and standard deviation, 24.27 ± 1.91) participated in Experiment 1. There were 18 participants (9 male; age range, 18–22 years; mean and standard deviation, 19.78 ± 1.17) in Experiment 2, 19 (9 male; age range, 18–21 years; mean and standard deviation, 19.47 ± 0.91) in Experiment 3, and 20 (6 male; age range, 18–31 years; mean and standard deviation, 23.85 ± 3.22) in Experiment 4. All participants were right-handed and had normal or corrected-to-normal vision. Consent forms were provided before the experiments according to the local University ethics committee.
Stimuli and task
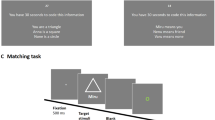
Participants were told to form associations between three types of geometric shapes (ellipses, rectangles, and triangles)Footnote 1 and three people (self, mother, and stranger). For example, the rectangles might represent a participant’s mother, the triangles might represent the participant (“you”), and the ellipses might represent an unfamiliar other (“stranger”). After the associative instruction, participants carried out a shape–label matching task where a shape preceded a label (‘mother,” “you,” or “stranger”) presented in the center of the screen. Sui, He, and Humphreys (2012) showed that there were minimal effects of variations in word length on shape–label matching. The shape–label pair either matched (e.g., mother–rectangle) or mismatched (e.g., mother–triangle). Participants had to judge whether the pair matched or mismatched. The shapes themselves could be large or small (~7˚ × 7˚ or ~3˚ × 3˚ of visual angle). Previous studies have used only a single shape–label pair (Sui, He, & Humphreys, 2012; Sui et al., 2013). Here, we used up to 40 different shapes for each matching label (rectangle, ellipses, and triangles varying in their dimensions) and two different overall sizes per shape in order to test matching at a more conceptual level than with a single shape instance. There was a set of 40 stimuli for each type of shape—half presented at a large size and half at a small size. These stimuli were randomly chosen for each display. A label subtended ~3˚ × 1.5 ˚ of visual angle. A star of 0.58° × 0.58° served for central fixation and was presented at the beginning of each trial. All the stimuli were displayed in white against a dark gray background. The assignment of shapes was counterbalanced across participants. The experiment was run on a PC using E-Prime software (Version 1.1). The stimuli were displayed on a 21-in. monitor (1,024 × 768 at 60 Hz).
The probability of the three matched shape–label pairs was manipulated across four experiments. There was an equal number of trials for each type of association in Experiment 1 (the baseline experiment). For Experiment 2, the probability for self-associated trials was reduced so that the ratio of the self, mother, and stranger match trials was approximately 1:3:3 (i.e., self matches occurred on 6 % of the trials, and mother and stranger matches occurred on 22 % of the trials; the other 50 % of the trials were mismatched). In Experiment 3, the probability for mother association match trials was reduced (self, mother, and stranger = 3:1:3 [22 %, 6 %, and 22 % of the trials]). In Experiment 4, there was a reduction for the stranger association match trials (self, mother, and stranger = 3:3:1 [22 %, 22 %, and 6 % of the trials]).
Procedure
For Experiments 1–4, a trial began with a central fixation for 500 ms, followed by a centrally presented shape for 100 ms. After a 200-ms blank interval, a label (“mother,” “you,” or “stranger”) appeared in the center of screen for 100 ms. Participants had to make a judgment as to whether the pairing of the shape and label matched within a variable response interval of 1,000–1,300 ms, during which a blank screen was presented. The response deadline was adopted in order to encourage fast responses and discourage strategic decision making. Participants were instructed to make a response by pressing one of the two keys on a keyboard as accurately and quickly as possible. Feedback was given for 500 ms once a response was made, and then the next trial started. Participants performed six blocks of 60 trials. In each block, the different experimental conditions were randomly presented. There were 18 trials for practice before each experiment began. There were 60 trials for each type of matching and mismatching conditions in Experiment 1. Mismatching trials were defined by the shape that was present. There were 30 trials for each shape re-paired with one of two mismatched labels (e.g., 30 trials for the self-associated shape re-paired with the “mother” label and 30 trials for the self-associated shape re-paired with the “stranger” label). There were 24 trials for the low-probability matching associations in Experiments 2–4 and 78 trials for the high-probability associations.
Data analysis
The trials with large and small shapes were merged because preanalyses showed no significant effects of shape size and no interactions involving shape size and shape–label association for any experiment. Separate data analyses were conducted for the matching and shape-based mismatching trials, partly because of the different response strategies that could be used and also to test the probability effect for matched trials. There was no trade-off between reaction times (RTs) and accuracy over the experimental conditions. The RT data are presented graphically, while the mean errors are tabulated (Table 1).
We report the results in three parts. First, a repeated measures analysis of variance (ANOVA) was conducted on the match trials in each experiment with a single within-subjects variable—association (self, mother, or stranger). Second, cross-experimental analyses with the same within-subjects factor of association (self, mother, or stranger) and a between-subjects factor of experiment (contrasting each of Experiments 2, 3, and 4 with the baseline Experiment 1) were then carried out in order to test how altering the probability of the different match trials affected performance. Finally, the effect of person–shape associations for shape-based mismatch trials was examined using a single ANOVA with a within-subjects factor of association (sorted according to whether the shape related to the self, mother, or stranger) and a between-subjects factor of experiment (Experiments 1–4; note that mismatch trials were identical across the experiments). Holm–Bonferroni corrections for α = .05 were applied to all multiple comparisons (Holm, 1979).
Results
Biases within each experiment
Experiment 1 (equal probabilities)
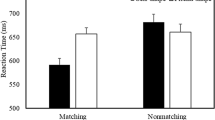
ANOVA on correct RTs revealed a reliable effect of association for matched pairs, F(2, 42) = 26.62, p < .001, η 2 = .56. Paired sample t tests showed that responses were faster to the self, as compared with the mother, t(21) = −3.17, p = .005, and stranger, t(21) = −8.43, p < .001, associations, and for the mother, as compared with the stranger, association, t(21) = −3.75, p = .001 (Fig. 1a).

Mean reaction times for matched trials in Experiments 1–4 as a function of association (self, mother, or stranger), in Experiment 1 with equal number of trials per condition (a), in Experiment 2 with reduction of probability for the self association (b), in Experiment 3 with reduction of probability for the mother association (c), and in Experiment 4 with reduction of probability for the stranger association (d). Error bars represent standard errors
An equivalent ANOVA on errors revealed a reliable effect of association for matched pairs, F(2, 42) = 14.88, p < .001, η 2 = .42. There were more errors to the stranger than to the self, t(21) = 4.04, p = .001, and mother, t(21) = 4.01, p = .001, associations, while there was no difference between the self and mother associations, t(21) = 0.98, p = .34 (Table 1).
Experiment 2 (low probability self matches)
For match trials, the analysis showed a significant effect of association, F(2, 34) = 15.01, p < .001, η 2 = .47. There were faster responses to the self and mother associations than to stranger association, t(17) = −5.69 and −4.80, ps < .001, but no significant difference between the self and mother associations was observed, t(21) = −0.76, p = .46 (Fig. 1b).
The ANOVA on errors did not show a significant effect of association, F(2, 34) = 1.16, p = .33 (Table 1).
Experiment 3 (low-probability mother matches)
The analysis on correct RTs demonstrated a significant effect of association for matched pairs, F(2, 36) = 81.77, p < .001, η 2 = .82. The result was due to responses being faster for match trials associated with the self, as compared with mother and stranger association trials, t(18) = −9.55 and −15.16, ps < .001); there was no difference between mother- and stranger-associated trial shapes, t(18) = 0.48, p = .64 (Fig. 1c).
The ANOVA on errors revealed a reliable effect of association for matched pairs, F(2, 36) = 14.10, p < .001, η 2 = .44. There were more error responses to the mother, t(18) = 5.11, p < .001, and stranger, t(18) = 4.83, p < .001, stimuli than to self-associated stimuli; but there was no difference between the mother and the stranger associations, t(18) = 0.45, p = 0.66 (Table 1).
Experiment 4 (low-probability stranger matches)
The RT analysis for the matched pairs demonstrated a significant effect of association, F(2, 38) = 80.13, p < .001, η 2 = .81. RTs were shorter to matched self than to matched mother and stranger associations, t(19) = −5.74 and −11.37, ps < .001; RTs were also shorter to mother- than to stranger-associated shapes, t(19) = −2.32, p < .001 (Fig. 1d).
The analysis on errors showed a significant main effect of association for matched pairs, F(2, 38) = 11.55, p < .001, η 2 = .38. There were more accurate responses to the self than to mother, t(19) = −2.81, p = .011, and stranger, t(19) = −4.91, p < .001, associations; there were also more accurate responses to the mother than to the stranger associations, t(19) = −2.32, p = .032 (Table 1).
Cross-experimental comparisons with baseline
In order to test how variations in the probability of the different match trials affected performance, cross-experimental analyses were performed relative to Experiment 1 (the baseline).
Effect of reduced probability for matching pairs on RTs
For the comparison of Experiments 1 and 2 (reduced probability for matched self pairs), there were significant main effects of association, F(2, 76) = 37.99, p < .001, η 2 = .50, and experiment, F(1, 38) = 7.23, p < .02, η 2 = .16. There was also a significant interaction, F(2, 76) = 3.41, p < .04, η 2 = .08. For both the mother and stranger associations, response latencies decreased in Experiment 2, as compared with Experiment 1, t(38) = −2.74 and −3.18, p = .009 and .003, reflecting the relatively higher probabilities of these trial types. However, reducing the probability of self matches did not affect response latencies to self-associated stimuli, t(38) = −1.37, p = .18 (Fig. 1a, b).
The comparison between Experiments 1 and 3 (reduced probability of mother matching pairs) showed a significant main effect of association, F(2, 78) = 85.46, p < .001, η 2 = .69, but no significant main effect of experiment, p = .31. The interaction between association and experiment was significant, F(2, 78) = 10.89, p < .001, η 2 = .22. The latency of responses to matched self trials decreased in Experiment 3, as compared with Experiment 1, t(39) = −2.64, p = .012, while there was no effect on matched trials for mother and stranger stimuli, t(39) = 0.85 and −1.43, p = .40 and .16 (Fig. 1a, c).
The analysis contrasting Experiments 1 and 4 (reduced probability of stranger matching pairs) showed a significant effect of association, F(2, 80) = 102.32, p < .001, η 2 = .72, but no effect of experiment, p = .73. There was a significant interaction between association and experiment, F(2, 80) = 10.54, p < .001, η 2 = .21. There was a tendency for response latencies to matched self shapes to decrease in Experiment 4, as compared with Experiment 1, t(40) = −1.87, p = .069, but there were no significant differences on mother and stranger association trials between Experiments 4 and 1, t(40) = −0.78 and 1.43, p = .44 and .16 (Fig. 1a, d).
Effect of reduced probability for matching pairs on error responses
The comparison of Experiments 1 and 2 (reduced probability for matched self pairs) revealed a significant main effect of association, F(2, 76) = 12.67, p < .001, η 2 = .25, but no significant effect of experiment, p = .74. The interaction between association and experiment was also significant, F(2, 76) = 5.03, p < .01, η 2 = .12. Post hoc t tests failed to show any significant differences between Experiments 1 and 2 for any of the associations [self, t(38) = −1.17, p = .25; mother, t(38) = −0.84, p = .41; stranger, t(38) = 1.89, p = .07].
For the comparison between Experiments 1 and 3 (reduced probability of mother matching pairs), there was a significant main effect of association, F(2, 78) = 21.83, p < .001, η 2 = .36, and no significant main effect of experiment, p = .30. The interaction between association and experiment was significant, F(2, 78) = 7.42, p = .001, η 2 = .16. Error responses for matched mother pairs increased when the probability of these trials was reduced in Experiment 3, as compared with Experiment 1, t(39) = 3.55, p = .001. In contrast, there were no effects on matched self, t(39) = −1.52, p = .14, and stranger, t(39) = −0.44, p = .66, trials.
The comparison contrasting Experiments 1 and 4 showed a significant effect of association, F(2, 80) = 23.92, p < .001, η 2 = .37; there were more accurate responses to the self than to the mother and stranger associations, t(41) = −2.82 and −6.17, p = .007 and p < .001, and more accurate responses to the mother than to the stranger associations, t(41) = −3.91, p < .001. Neither the main effect of experiment nor the interaction between association and experiment was significant (p = .10 and .15).
Effect of increasing the probability of matching pairs on RTs
In order to test how increases in the frequency of occurrence for matching pairs modulated performance, we conducted a comparison between Experiment 1 (equal stimulus probabilities) and Experiments 3 and 4 (increased probability of occurrence for matched self pairs) for self and other pairs separately. An independent samples test for self pairs revealed that responses were faster when the probability of self matches increased in Experiments 3 and 4 (mean and SE: 514 ± 11), as compared with Experiment 1 (mean and SE: 570 ± 19), t(59) = −2.71, p = .009. In contrast, increases in the proportions of familiar “other” pairs (stranger or mother, respectively, in Experiments 3 and 4; mean and SE: 684 ± 17) did not facilitate the latency of responses, as compared with Experiment 1 (mean and SE across the mother and stranger trials: 648 ± 18), t(59) = 1.33, p = .19.
Effect of increasing the probability for matching pairs on error responses
The analysis of error performance revealed no significant difference in the self association condition when the probability of self match trials increased in Experiments 3 and 4 (mean and SE: 0.02 ± 0.003), as compared with Experiment 1 (mean and SE: 0.03 ± 0.010), p = .094. However there were more error responses when the probability of familiar “other” pairs increased in Experiments 3 and 4 (mean and SE: 0.16 ± 0.010), as compared with Experiment 1 (mean and SE: 0.09 ± 0.01), t(59) = 2.38, p = .020.
Analysis for mismatched pairs
RTs and errors for mismatching pairs, presented as a function of the shape on the mismatching trial, are shown in Tables 1 and 2. The ANOVA on RTs revealed a significant effect of association, F(2, 150) = 26.46, p < .001, η 2 = .26, due to faster responses to self- than to mother- and stranger-associated shapes, t(78) = −5.23 and −6.59, ps < .001, while there was no difference between the mother- and stranger-associated shapes, t(78) = −1.42, p = .16 (Fig. 2). The effect of experiment was marginal, F(3, 75) = 2.54, p = .06, η 2 = .09, and no significant interaction between association and experiment was observed, F(6, 150) = 1.05, p = .39.

Means of reaction times for shape-based mismatched trials across four experiments as a function of association (self, mother, or stranger). Error bars represent standard errors
The ANOVA on error performance showed a significant effect of association, F(2, 150) = 4.50, p < .02, η 2 = .06, reflecting more accurate responses to the self- than to mother- and stranger- associated shapes, t(78) = −2.74 and −2.38, p = .008 and .020, but there was no difference between the mother- and stranger-associated shapes, t(78) = 0.83, p = .41. There was no significant main effect of experiment, and the interaction between association and experiment was not reliably significant, F(6, 150) = 0.51 and 1.57, p = .68 and .16.
Discussion
When compared with the stranger condition, there was a self-bias effect in all experiments, even when the self match was a low-probability event (and when there was a high probability of a mismatch trial following a self shape, as compared with a match trial, and so, participants should have actively predicted a mismatch response; cf. Schmidt & Besner, 2008). In contrast, the mother advantage (relative to stranger match trials) was eliminated when mother match trials were low-probability events. The data point to the self advantage on match trials being highly robust and relatively immune to manipulations of the probability of the event.
This last conclusion is supported by the across-experiment comparison, which revealed that, relative to the baseline experiment with equal probabilities of match trials (Experiment 1), performance on self trials was unaffected by making self matches relatively low-probability events. In contrast, for example, mother match trials were made relatively less efficient when these trials were lower probability events. The failure to lessen the self advantage when self match trials occurred with a low probability contrasts with the results on prepotent responses such as word naming in experiments on the Stroop effect and location-based responding in experiments on the Simon effect (e.g., Logan et al., 1984; Milliken & Lupiáñez, 2005; Schmidt, 2013; Schouppe, De Houwer, Ridderinkhof, & Notebaert, 2012; Stürmer et al., 2002), since these effects are reduced when congruent trials decrease. The results highlight the continued potency of the self over and above the effects of probability. In addition, the contrast between the robust self advantage across different probabilities and the decreasing advantage for mother trials when matching stimuli decreased in the mother condition (Experiment 3) provides evidence that the self advantage effect on associative shape matching is not simply driven by the familiarity of self-related stimuli. Previous work has shown that there are close relations between self and mother representations for Chinese participants, even at a neural level (Wang et al., 2012; Zhu et al., 2007), suggesting that mother representations are closer to the self than to representations of strangers, for individuals in this culture. We used Chinese participants, and yet we still demonstrated contrasting effects of probability for self- and mother-associated stimuli. These results indicate that, at the very least, the self advantage does not represent a simply linear shift in familiarity (since the mother advantage was eliminated and the self advantage maintained, relative to stranger trials), and indeed, the results are consistent with the self advantage here not being based on differential familiarity at all. Of course, this leaves unresolved exactly which factors are critical. Sui, He, and Humphreys (2012) showed that effects of high reward generated advantages similar to those of the self on associative shape matching, and so it is possible that the self advantage is mediated by high intrinsic reward values linked to the self (Northoff & Hayes, 2011). An alternative possibility is that self-associated stimuli elicit positive emotional responses that facilitate matching performance (Ma & Han, 2010). Whether high-reward and positive-emotion linked stimuli are robust to variations in their probability of occurrence is a question for future research.
Although there was a robust self advantage that was the same magnitude when the self match trials occurred with probabilities that were reduced from or equal to those of other types of trial, this advantage did increase when the self match trials increased in their relative probability (in Experiments 3 and 4, as compared with Experiment 1). This is consistent with there being two components to the self bias: (1) a component that appears to be relatively automatic and immune to the likelihood of stimulus events, which is present even when self matches occur with a low probability (Experiment 2), and (2) an effect that increases when self matches are relatively high-probability events (Experiments 3 and 4). As was noted in the Introduction, there is controversy about the factors that modulate stimulus probability effects under Stroop and Simon conditions. On the one hand, some have argued for effects of expectancy (Gratton et al., 1992), while others have argued for implicit learning effects (Blais et al., 2012), which themselves might arise in a relatively automatic fashion (Schmidt & De Houwer, 2011). Although we did not specifically aim to test the mechanisms by which stimulus likelihood affects performance, the present results are relevant to this debate. Notably, in the experiments where self matches had a higher probability of occurring, there were minimal increases in performance for the other higher probability conditions (the stranger in Experiment 3 and the mother in Experiment 4), even though they had the same likelihood of occurrence as self trials. In contrast, in Experiment 2, when mother and stranger trials were both high probability and self trials were infrequent, there were benefits for both mother and stranger match conditions. This last result indicates that there was no inherent limitation on performance benefits to two stimulus types when each occurred with a higher probability. It is difficult to understand these variations in performance for self-related stimuli, on the one hand, and stimuli related to other people, on the other hand, simply in terms of stimulus–response contingency learning, since stimulus–response contingencies were the same for the different stimuli. However, the results can be accounted for if participants employed specific expectancies based on stimulus probabilities and if these expectancies favor self-related stimuli over stimuli associated with other people. The data indicate that, when self-related stimuli were expected, participants were biased to the self over and above other stimuli (eliminating the benefit for high-frequency stranger trials in Experiment 3 and for high-frequency mother trials in Experiment 4). Apparently, having an expectancy for self-related stimuli operates to the exclusion of holding expectancies for stimuli related to other people. In contrast, when the mother- and stranger-associated stimuli both had a relatively high probability of occurrence, both benefitted (Experiment 2 vs. Experiment 1). There has been much recent work indicating that effects of expectancy on visual perception and attention are mediated by holding the expected stimuli in working memory (see Soto, Hodsoll, Rotshtein, & Humphreys, 2008, for one review). Furthermore, it has been argued that improvements in processing are particularly strong when stimuli match representations held at the “forefront” of working memory (see Olivers, 2009), as compared with when stimuli are represented more in the background. Here, we may speculate that self-related expectancies are maintained at the forefront of working memory, “pushing” expectancies for other high-probability events to the background. This generates strong facilitation effects for self trials and minimizes the benefits for other high-probability events. With expectancies for stimuli related to other people, either both expectancies may be held (but perhaps more weakly) in the foreground of working memory or only one is held (Olivers, 2009), but this alternates over trials. The net result is that benefits can then emerge for both associations.
Finally, we note two other, more minor aspects of the data. One is that the present results occurred even though shape–label matches were not performed to single shape instances but, rather, to multiple instances of the same type of shape. That is, matching here was more likely to operate at a conceptual rather than a low-level perceptual level. There is a strong self bias both when single (Sui, He, & Humphreys, 2012; Sui et al., 2013) and when multiple shape instances are used here.
In addition, there were quicker mismatch responses following a self shape as the cue on a trial, as compared with when the initial stimulus was a shape for another person, and this was particularly the case in Experiment 2. This is interesting because these faster mismatch responses fit with there being a conscious expectation for a mismatch event after a self shape when self match trials were infrequent (and self mismatch trials were more frequent events). There remained a robust self advantage on match trials, however (as compared with the stranger match baseline). This is consistent with the automatic component of the self match advantage being present and influencing self match responses, even though the expectation was for a mismatch response (cf. Schmidt & Besner, 2008).
Conclusions
In sum, the study provides robust evidence on how personal significance (for self-associated stimuli, as compared with stimuli associated to a very close other, the mother) impacts on perceptual matching. The results indicate that self biases can emerge in at least two ways: from automatic bottom-up biases that are impervious to the effects of stimulus probability and from what we suggest are top-down expectancies, used when stimuli have a high probability of occurrence. The data provide a new perspective on how social associations can shape our cognition and behaviors.
Notes
A control experiment was conducted showing no differences in shape perception between the different shapes, indicating no systematic biases introduced by the shapes used.
References
Blais, C., Harris, M. B., Guerrero, J. V., & Bunge, S. A. (2012). Rethinking the role of automaticity in cognitive control. Quarterly Journal of Experimental Psychology, 65, 268–276.
Brédart, S., Delchambre, M., & Laureys, S. (2006). One’s own face is hard to ignore. The Quarterly Journal of Experimental Psychology, 59, 46–52.
Gratton, G., Coles, M. G. H., & Donchin, E. (1992). Optimizing the use of information: Strategic control of activation of responses. Journal of Experimental Psychology: General, 121, 480–506.
Holm, S. (1979). A simple sequentially rejective multiple test procedure. Scandinavian Journal of Statistics, 6, 65–70.
Kumar, S., Yoon, E. Y., & Humphreys, G. W. (2012). Perceptual and motor-based responses to hand actions on objects: Evidence from ERPs. Experimental Brain Research, 220, 153–164.
Logan, G. D., Zbrodoff, N. J., & Willliamson, J. (1984). Strategies in the color-word Stroop task. Bulletin of the Psychonomic Society, 22, 135–138.
Ma, Y., & Han, S. (2010). Why respond faster to the self than others? An implicit positive association theory of self advantage during implicit face recognition. Journal of Experimental Psychology: Human Perception and Performance, 36, 619–633.
MacLeod, C. M. (1991). Half a century of research on the Stroop effect: An integrative review. Psychological Bulletin, 109, 163–203.
Milliken, B., & Lupiáñez, J. (2005). Repetition costs in word identification: Evaluating a stimulus-response integration account. Psychology Research, 71, 64–76.
Northoff, G., & Hayes, D. J. (2011). Is our self nothing but reward? Biological Psychiatry, 69, 1019–1025.
Olivers, C. N. L. (2009). What drives memory-driven attentional capture? The effects of memory type, display type, and search type. Journal of Experimental Psychology: Human Perception and Performance, 35, 1275–1291.
Schmidt, J. R. (2013). Questioning conflicting adaptation: Proportion congruent and Gratton effects reconsidered. Psychonomic Bulletin and Review, 20, 615–630.
Schmidt, J. R., & Besner, D. (2008). The Stroop effect: Why proportion congruent has nothing to do with congruency and everything to do with contingency. Journal of Experimental Psychology: Learning, Memory, and Cognition, 34, 514–523.
Schmidt, J. R., & De Houwer, J. (2011). Now you see it, now you don’t: Controlling for contingencies and stimulus repetitions eliminates the Gratton effect. Acta Psychologica, 138, 176–186.
Schmidt, J. R., De Houwer, J., & Besner, D. (2010). Contingency learning in the blink of an eye: A resource dependent process. Consciousness and Cognition, 19, 235–250.
Schouppe, N., De Houwer, J., Ridderinkhof, K. R., & Notebaert, W. (2012). Conflict: Run! Reduced Stroop interference with avoidance responses. Quarterly Journal of Experimental Psychology, 65, 1052–1058.
Simon, J. R. (1969). Reactions towards the source of stimulation. Journal of Experimental Psychology, 81, 174–176.
Soto, D., Hodsoll, J. P., Rotshtein, P., & Humphreys, G. W. (2008). Automatic guidance of attention from working memory. Trends in Cognitive Science, 12, 342–348.
Stroop, J. R. (1935). Studies of interference in serial verbal reactions. Journal of Experimental Psychology, 18, 643–662.
Stürmer, B., Leuthold, H., Soetens, E., Schröter, H., & Sommer, W. (2002). Control over location-based response activation in the Simon task: Behavioral and electrophysiological evidence. Journal of Experimental Psychology: Human Perception and Performance, 28, 1345–1363.
Sui, J., Chechlacz, M., & Humphreys, G. W. (2012). Dividing the self: Distinct neural substrates of task-based and automatic self-prioritization after brain damage. Cognition, 122, 150–162.
Sui, J., He, X., & Humphreys, G. W. (2012). Perceptual effects of social salience: Evidence from self-prioritization effects on perceptual matching. Journal of Experimental Psychology: Human Perception and Performance, 38, 1105–1117.
Sui, J., Rotshtein, P., & Humphreys, G. W. (2013). Coupling social attention to the self forms a network for personal significance. Proceedings of the National Academy of Sciences of the United States of America, 110, 7607–7612.
Wang, G., Mao, L., Ma, Y., Yang, X., Cao, J., Liu, X., ... Han, S. (2012). Neural representations of close others in collectivistic brains. Social Cognitive and Affective Neuroscience, 7, 222–229.
Yoon, E. Y., & Humphreys, G. W. (2005). Direct and indirect effects of action on object classification. Memory & Cognition, 33, 1131–1146.
Zhu, Y., & Zhang, L. (2002). An experimental study on the self-reference effect. Science in China Series C, 45, 120–128.
Zhu, Y., Zhang, L., Fan, J., & Han, S. (2007). Neural basis of cultural influence on self-representation. NeuroImage, 34, 1310–1316.
Acknowledgments
This work was supported by the National Nature Science Foundation of China (Project 31371017) and by grants from the Economic and Social Research Council (ES/J001597/1, UK) and the European Research Council (Pepe Grant 323883).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Sui, J., Sun, Y., Peng, K. et al. The automatic and the expected self: separating self- and familiarity biases effects by manipulating stimulus probability. Atten Percept Psychophys 76, 1176–1184 (2014). https://doi.org/10.3758/s13414-014-0631-5
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13414-014-0631-5