
Abstract
Attention shifts are facilitated if the items to be attended remain the same across trials. Some researchers argue that this priming effect is perceptual, whereas others propose that priming is postperceptual, involving facilitated response selection. The experimental findings have not been consistent regarding the roles of variables such as task difficulty, response repetition, expectancies, and decision-making. Position priming, when repetition of a target position facilitates responses on a subsequent trial, is another source of disagreement among researchers. Experimental results have likewise been inconsistent as to whether position priming is dependent on the repetition of target features or has an independent effect on attention shifts. We attempted to isolate the perceptual components of priming by presenting brief (10–180 ms) search arrays to eight healthy observers. The task was to identify a color-singleton letter among distractors. All stimulus presentation contingencies were randomized, and responses were unspeeded, to avoid effects of observer expectation and postperceptual effects. Repeating target color and/or position strongly improved performance. The effects of color and position repetition were independent of one another and were stable across participants. The results argue for a strong perceptual component in priming, which biases selection toward recent target features and positions, showing that perceptual mechanisms are sufficient to produce priming in visual search and that such effects can be elicited with limited sensory evidence. The results are the first to demonstrate independent priming of color and position in the identification of briefly presented, postmasked stimuli.
Similar content being viewed by others
When searching the visual environment, whether for favorite pieces in a bowl of candy or when seeking fungi in the forest undergrowth, observers repeatedly search for similar visual features. Biasing of selective attention toward recently important visual features, or away from recently dismissed ones, therefore carries obvious benefits. In laboratory studies designed to test such behavior in controlled settings, observers repeatedly search for a target of uncertain identity. These studies have revealed numerous examples of repetition priming, in which performance improves as targets are repeated on consecutive trials. The priming effects typically appear as reaction time benefits when a target identity is repeated, as compared with when it changes. The literature on priming suggests that visually similar real-world objects will be found more quickly when they are searched for on successive trials, but also that observers will show a preference for stimuli similar to a recent target, given free choice between two equally relevant ones (Brascamp, Blake, & Kristjánsson, 2011), a behavior analogous to foraging (Dawkins, 1971).
This facilitated responding to recently attended objects, as well as the statistical preference for those objects during free choice, could be accounted for in a number of plausible ways. These differ in what mechanism is assumed to be the carrier of the repetition facilitation, ranging from simple visual features to complex interactions between multiple features and decision-making.
The simplest proposal is passive short-term potentiation of neurons sensitive to repeated, and therefore primed, features—for example, “red” or “vertical.” Such accounts entail that priming facilitates responses to targets with particular features, regardless of whether the features are attended or not. Comparing tasks that do and do not involve attentional selection allows for testing whether sensory potentiation is responsible for facilitated responses on trials with repeated target features. Goolsby and Suzuki (2001) tested observers on a conventional pop-out search task, as well as on a similar task with exogenous precues about upcoming target position. Precuing makes selection trivial, since observers can allocate attention to a specific position before any stimuli appear. On precued trials, color repetition effects were negligible, whereas strong priming effects emerged from noncued trials (Goolsby & Suzuki, 2001). This result is incompatible with a passive-potentiation account for priming (see also Kristjánsson, Saevarsson, & Driver, 2013). That priming requires a selection criterion is corroborated by findings from whole-report tasks (see Sperling, 1960); when all briefly presented visual elements are equally relevant, no benefit of color repetition arises (Ásgeirsson, Kyllingsbæk, Kristjánsson, & Bundesen, 2012).
A more viable explanation of priming involves feature facilitation, in which different features compete for processing, and those of recent importance are prioritized. Multiple versions of such accounts exist (Becker & Horstmann, 2009; Kristjánsson, 2006a; Maljkovic & Nakayama, 1994; see Kristjánsson, 2008, for review). What these accounts have in common is that, first, they predict priming in conditions in which selective attention is needed, and second, the priming effect is driven by the repetition of a feature, rather than of a conjunction of features. Importantly, postperceptual processes, such as decision-making or response mapping, play no role in such accounts.
In addition to the more conventional reaction time benefits, feature-facilitation accounts of priming predict target repetition benefits in accuracy tasks through increased selection efficiency of repeated features. Furthermore, they predict that primed features will be processed independently and that repetition benefits from different features should not interact (Kristjánsson, 2006b, 2009; Maljkovic & Nakayama, 1994, 1996).
Perceptual priming has also been found for whole objects, in particular for stimuli particularly susceptible to object processing (Kristjánsson, Ingvarsdóttir, & Teitsdóttir, 2008). Kristjánsson et al. (2008) explored whether priming would differ by stimulus type. In one experiment, observers looked for an odd-one-out diamond made up of two colors, divided by a vertical line, whereas in another experiment the diamonds were made up of a small square enclosed within a larger one. Enclosure is commonly associated with object-based processing (e.g., Vogel, Woodman, & Luck, 2001; Wolfe, Friedman-Hill, & Bilsky, 1994). In both experiments, the task was to identify the odd-diamond-out and report whether a notch had been cut from its top or bottom corner. Three conditions were compared, in which (1) neither target color repeated, (2) one target color repeated, and (3) both colors repeated. In the first experiment, a linear benefit of repetition increased from no to one to two color repetitions, whereas in the second experiment, priming was only seen when both colors repeated. This suggests that the primed unit (feature or feature conjunction) depends on the processing demands of the stimulus.
According to other accounts, however, priming reflects postperceptual biases of decision-making or response facilitation (Hillstrom, 2000; Huang, Holcombe, & Pashler, 2004), Such accounts do not predict priming benefits in pure accuracy tasks; if there is no pressure to respond quickly, decisions can be made, and motor responses chosen, with care. Huang and Pashler (2005) tested repetition benefits in a pure accuracy visual search task and found no benefits of feature repetition, unless the presentation contingencies were such that the observers’ expectations for a particular target were heightened. They concluded that expectancies affect perception, but that “[t]his perceptual improvement cannot be fully explained by either target-feature enhancement or distractor-feature inhibition, nor by their summation” (Huang & Pashler, 2005, p. 157). They further concluded that the well-known priming of pop-out effect (Maljkovic & Nakayama, 1994) is exclusively postperceptual. This falls in line with other claims by the same authors (Huang et al., 2004) that priming is postperceptual, is based on episodic memory retrieval, and involves mapping a particular response to a visual object. But whereas the first study failed to demonstrate priming effects in the absence of repetition expectancy, and the latter demonstrated maximal priming when two feature repetitions coincided with response repetitions, the results do not generalize well to slight variations of the originally tested task (Ásgeirsson & Kristjánsson, 2011; see also Sigurdardottir, Kristjánsson, & Driver, 2008; Yashar & Lamy, 2010).
Two studies are particularly important, regarding the question of whether priming can occur in isolation from postperceptual processes. Sigurdardottir et al. (2008) showed that repetition streaks increased perceptual sensitivity to color singletons in brief (200-ms) exposure search tasks. It is, however, possible that the effects were partly due to learning of statistical contingencies, since target and distractor identity did not vary randomly (cf. Huang & Pashler, 2005). Repetitions followed a probabilistic rule intended to increase repetition likelihood: Was the increased perceptual sensitivity due to feature facilitation or to expectation based on predictability, as Huang and Pashler (2005) would predict? The distinction is important, since one alternative involves a short-term memory mechanism (with a capacity of a few trials; Nakayama, Maljkovic, & Kristjánsson, 2004), whereas the other involves a longer-term memory mechanism that gathers information about task contingencies.
Yashar and Lamy (2010) ruled expectancy out by varying the defining (shape) and response (orientation of a “T” embedded within the shape) variables randomly in a brief exposure visual search task. Participants were either to allocate focal attention to a shape singleton or to make a coarse judgment of which hemifield a target was presented in. Priming effects only arose in the former condition. The authors concluded that for perceptual-priming effects, focal attention must be deployed to the target during encoding, and again during retrieval of the primed target, explaining the discrepant results from previous studies (i.e., Huang & Pashler, 2005, and Sigurdardottir et al., 2008).
The question of whether target feature and position repetition interact has received less attention than have interdependencies of visual features and response demands (Hillstrom, 2000) or of visual features with different task relevance (Kristjánsson, 2006b, 2009). Spatial repetition may produce the most complex and least predictable priming effects. They are complicated by phenomena such as inhibition of return (IOR; see Wang & Klein, 2010, for a recent review) and were initially only observed as co-occurring with feature repetition (Treisman, Vieira, & Hayes, 1992). In the first study specifically designed to explore and measure position priming (Maljkovic & Nakayama, 1996), the effect did not depend on simultaneous feature repetition, but the largest position effects stemmed from the target before the most recent one. The authors suggested that this might have been caused by weak IOR modulation of strong position-priming effects.
Campana and Casco (2009) investigated which conditions result in facilitation or inhibition of return in visual search. They compared two task types, a feature and a position discrimination task. Their results suggested that feature repetition was always facilitatory, whereas position repetition effects were task-dependent. When position was related to the motor response, an inhibitory effect of repeating position emerged when the target feature changed (color or orientation), but a benefit arose when both position and feature repeated. However, when position was task-irrelevant, position priming occurred along with feature priming, but did not affect trials on which the target feature changed between trials (see also Pratt & Castell, 2001). This suggests a relatively complex task-dependent interplay between implicit memory mechanisms. Campana and Casco suggested that spatial repetition effects reflected the operation of different mechanisms than did feature repetition, perhaps in areas in which spatiomotor transformations are processed (Campana & Casco, 2009, p. 336). According to this view, position priming is a postperceptual effect involving the mapping of a stimulus to an appropriate response (see Huang et al., 2004, for a related account of feature priming), rather than a consequence of selective visual attention. Contrast this view of position priming with Yashar and Lamy’s (2010) conclusion that feature priming is primarily a “selection phenomenon,” and the two types of repetition effects (position and feature priming) seem much less heterogeneous than earlier interpretations had suggested (Maljkovic & Nakayama, 1996).
Present aims
Our first goal was to test for priming effects from color and position repetition in a paradigm in which interpretations based on expectancy (Huang & Pashler, 2005) and postperceptual processes (i.e., Hillstrom, 2000; Huang et al., 2004) could be conclusively ruled out. We presented matrices of colored letters, in which the task was to report the oddly colored singleton. Second, we tested interactions between repetition of color and stimulus position. These attributes have sometimes been found to be independent (Maljkovic & Nakayama, 1996) and sometimes interdependent (Campana & Casco, 2009; Treisman et al., 1992) in conventional reaction time studies. Third, varying the exposure durations (10–180 ms) allowed us to investigate temporal aspects of priming, such as the minimum exposure time, that have not yet been addressed in the literature. Finally, we defined a rudimentary model in which we suggest an interpretation of the results within the framework of the theory of visual attention (TVA; Bundesen, 1990).
Method
Participants
The participants were eight University of Iceland students (five female, three male) 22–28 years of age, with normal or corrected-to-normal visual acuity (self-reported). Seven were naive volunteers, whereas one was a research assistant aware of the experimental hypotheses. All were native speakers of Icelandic.
Stimuli and apparatus
The stimuli were presented on a 19-in., 100-Hz CRT monitor controlled by a 2.4-GHz Dell desktop computer. Stimulus presentation was programmed in MATLAB using the Psychophysics Toolbox (version 3.0.9; Brainard, 1997; Pelli, 1997). Central fixation at the start of each trial was monitored monocularly at 60 Hz with a Cambridge Research Systems Video Eyetracker using the Video Eyetracking toolbox for MATLAB (Cambridge Research Systems, 2008).
The experimental stimuli were 15 uppercase consonants shared by the Icelandic and Latin alphabets (B, D, F, G, H, J, K, L, M, N, R, S, T, V, X). Character height was 3.0 deg of visual angle, and the font was Arial Bold. Target and distractor color varied randomly between a red target among blue distractors and the reverse combination (19 and 13 cd/m2, respectively). Pattern masks were made from crops of Arial Bold characters in the same red and blue. The stimulus attributes—colors, relative brightness, and masks—were adapted from the study of Vangkilde, Bundesen, and Coull (2011).
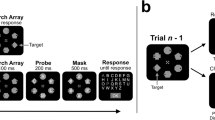
The stimuli were presented in matrices of 3 × 3 characters, in which each of the nine characters was unique (see Fig. 1). On each trial, one target defined by a unique color appeared among eight distractors (of the opposite color). To increase the number of position repetitions per participant, the target always occupied a corner position in the matrix, never the middle row or column. This ensured that the target was always flanked equally by nearby distractors. In sum, given all of the constraints that we have stated, the observer’s task was to search for a color singleton in one of two possible colors at four possible positions and report the letter identity of the singleton. Responses were made on a standard USB keyboard, in which any letter was reported by pressing the key matching the target letter identity.

Within-trial (solid arrow) and between-trial (dashed arrow) sequences. All trials started with a fixation period, during which the eyetracker waited for a 300-ms fixation. Following a brief delay, a matrix of 3 × 3 consonants appeared for 10–180 ms, followed by a 10-ms blank interval. The letter array was then masked for 500 ms, before a blank-screen period during which the observers were to make nonspeeded responses on a keyboard. The dashed arrow shows examples of stimulus variation between trials, in which trial n + 1 is a position repetition and n + 2 is a color repetition. The stimuli are not drawn to scale (see the Stimuli and apparatus section for the stimulus specifications)
Procedure and design
Experimental sessions were conducted in a dimly lit room with no direct lighting falling on the monitor or the observer. Before the experiment started, observers read illustrated task instructions. The observers’ eye movements were then calibrated, and they proceeded to a practice block of 50 trials.
Each trial started with a central fixation cross that remained onscreen until fixation (minimum of 300 ms) was confirmed. This was followed by a brief delay (100–300 ms), after which the fixation cross disappeared and the stimulus matrix appeared at screen center (the middle character replacing the fixation cross) for 10, 20, 30, 50, 70, or 180 ms (determined randomly). Subsequently, a blank interval of 10 ms was followed by 500-ms pattern masks. When the masks disappeared, a short beep (700-Hz sine wave) indicated that it was time to respond. The beep was played again as soon as the observer had responded with a keypress, and the next trial commenced. Recalibration of the eyetracker preceded every trial block.
Each observer participated in ten 100-trial blocks, totaling 1,000 trials. The conditions varied randomly and independently throughout the experiment, so that the prior probability of color repetition was always .5, whereas the probability of position repetition was .25. The prior probabilities for no repetition and color repetition, on the one hand, and for position repetition and repetitions of both color and position, on the other, were therefore .375 and .125, respectively. The actual values deviated from the prior probabilities by at most 2.7 percentage points for a single participant, whereas the minimum number of trials for a single condition was 107.
Results
To eliminate any effects of response repetition (see, e.g., Hillstrom, 2000), all trials on which target identity was the same as on the previous trial were discarded, leaving an average of 931 trials per observer. The average accuracy collapsed over all conditions and observers was 53 % (between-subjects range: 42 %–71 %). With so many incorrect trials, it is fair to assume that the exposure time was often too short to facilitate selection on subsequent trials. In fact, on some trials observers may not have seen the stimuli at all. Therefore, our analyses include only trials directly following correct responses. Our aim was to assess priming stemming from actually identified targets, rather than a mixture of potential and actual primes. The remaining data consisted of an average of 494 trials per observer (between-subjects range: 391–651).
Figure 2a shows mean accuracy as a function of exposure duration and color/position repetitions. A Color Repetition × Position Repetition × Exposure Duration repeated measures analysis of variance (ANOVA) revealed main effects of color repetition [6.0 percentage points; F(1, 7) = 25.18, RMSD1 = .089, p = .002, η 2 = .009] and position repetition [6.4 percentage points; F(1, 7) = 27.08, RMSD = .089, p = .001, η 2 = .01], but no interaction between the two [F(1, 7) = 0.09, RMSD = .110, p = .77, η 2 = .005]. Unsurprisingly, exposure duration was responsible for the largest chunk of variance in accuracy [F(5, 35) = 116.25, RMSD = .176, p < .001, η 2 = .858]. Neither color repetition nor position repetition interacted with exposure duration (Fs < 1.98, ps > .1).

(a) Data points show mean accuracy by exposure duration and repetition condition, collapsed across observers. The smooth curves show a plausible interpretation of the data, consistent with Bundesen’s (1990) theory of visual attention (TVA). According to this interpretation, the occurrence of a feature j (e.g., a color or a spatial position) as a feature of a target on trial n increases the pertinence of feature j on trial n + 1, so that more resources are allocated to processing objects appearing in the colors and spatial positions that were used for targets on the previous trial. The curves show least-squares fits by TVA, given three simplifying assumptions: (a) Visual confusions are neglected (see Kyllingsbaek, Markussen, & Bundesen, 2012); (b) selection by spatial location was perfect, such that only letters in the four corner positions of the stimulus matrix had positive attentional weights; and (c) visual short-term storage capacity was at least four letters. Given these simplifications, TVA predicts that P(correct) = 1 – exp[−v(t – t 0)], for t ≥ t 0, where processing rate v varies between the four conditions, t is the exposure duration, and t 0 is a perceptual threshold. The data are corrected for guessing by the standard high-threshold model (see Luce, 1963). The suggested model, based on the stated assumptions, explains 98.5 % of the variation in P(correct). (b) Probability-correct profiles of the individual observers (dotted lines) and the average profile across observers (thick black line)
The reported ANOVA on the observed response probabilities did not include a satisfactory test for independence between the effects of color repetition and position repetition. Independence between the effects of color and position repetition would imply that if, at a given exposure duration, the effect of color repetition was to multiply the probability p of a correct report by k 1, and the effect of position repetition was to multiply the probability p of a correct report by k 2, then the effect of color repetition combined with position repetition should be a multiplication of p by the product of k 1 and k 2. In terms of logarithms, we can say that if the effect of color repetition was to add log(k 1) to log(p), and the effect of position repetition was to add log(k 2) to log(p), then the effect of color repetition combined with position repetition should be an addition of log(k 1) + log(k 2) to log(p). Therefore, to test for independence between the effects of color and position repetition on the probability of a correct response, we tested for an additive relationship (i.e., lack of interaction) between the effects of color repetition and position repetition on the logarithms of the mean probabilities (collapsed over exposure durations). For the ANOVA on the log-transformed data, we again treated color repetition and position repetition as within-subjects factors in a repeated measures design. Both main effects, color [F(1, 7) = 21.72, RMSD = .071, p < .002, η 2 = .348] and position [F(1, 7) = 21.39, RMSD = .071, p = .002, η 2 = .345], were significant, whereas, importantly, we observed no interaction between color and position repetition [F(1, 7) = 0.015, RMSD = .063, p = .905, η 2 < .001]. The vanishingly low F value for the interaction term means that the test showed no signs at all that the results were due to a lack of statistical power. Thus, the results strongly suggest that the effects of color and position repetition are independent. Note that the raw probabilities for each observer are shown in Fig. 2b. The figure shows the exact data points that were log-transformed for the main ANOVA. The priming effects had fairly high stability across observers, in that six out of eight observers’ profiles adhered to a pattern of incremental priming with increasing repeated dimensions (from no, to one, to two dimensions repeated).
Maljkovic and Nakayama (1996) found a notable difference between position and feature priming (Maljkovic & Nakayama, 1994), in that the strongest priming from repeated features (color) came from the most recent trial, whereas position priming was strongest from two trials back (possibly due to IOR; see Wang & Klein, 2010, for a review). We tested this with a repeated measures ANOVA with the factors (n – 2) Position Repetition and Exposure Duration. We found a marginally significant main effect of (n – 2) position repetition [F(1, 7) = 5.013, RMSD = .071, p = .06, η 2 = .024] that was independent of exposure duration [F(2.67, 18.72)1 = 0.085, RMSD = .089, p = .957, η 2 < .001].Footnote 1
Finally, an exploratory repeated measures ANOVA of the subsets for each exposure duration showed that when the exposure duration was 20 ms, significant effects emerged of both color and position repetition (ps = .032 and .005, respectively). The p values for color repetition were below .05 for the 30- and 50-ms exposures, whereas the p value for the Position Repetition factor was only below the critical value at 20 and 70 ms. However, the p values derived from these analyses must be taken with a grain of salt, since they were not corrected for multiple comparisons. The comparisons are highly correlated, which makes a meaningful correction procedure difficult. The main message is that the descriptive data points form a coherent pattern, from the briefest exposure duration sufficient for selection until the observers reached ceiling performance (Fig. 2b).
Discussion
Our results show that priming plays a decisive role in perception when observers have to select an odd-one-out stimulus. The effects of repeating color and position were strong upon the accuracy of letter reports and replicated the previous findings of Sigurdardottir et al. (2008) and Yashar and Lamy (2010) in a novel task. Importantly, our results show, for the first time, independent effects of color and position repetition. This is reminiscent of the stochastic independence between the encoding of different dimensions that is assumed by many parallel-processing models (Bundesen, 1990; see also Bundesen, Kyllingsbæk, & Larsen, 2003, and Kyllingsbæk & Bundesen, 2007, for studies showing stochastic independence of processing of different stimulus dimensions). This is also the first study to show independent priming of features and position with briefly presented stimuli in conditions in which the artifacts of facilitated response selection, decision related processes, response repetition, and expectancies are conclusively ruled out.
The independence of position and feature priming may come as a surprise, in light of results showing interactions between these factors (Campana & Casco, 2009; Pratt & Castell, 2001; Treisman et al., 1992). Using reaction time measures, Campana and Casco observed significant interaction effects between feature and position repetition of about 100 ms (Cohen’s d > 0.5). We, however, found no meaningful difference in position priming dependent on feature repetition. This shows that selection benefits from target position repetition, independent of target identity or repetitions of other features. We may speculate that position-priming benefits, strong during target selection, disappear when postperceptual effects come into play in reaction-time-based search tasks. Given that these postperceptual effects depend on whether a target feature is repeated or changed (Campana & Casco, 2009), a plausible mechanism for these effects may be found at the stage of spatiomotor transformations (Campana & Casco, 2009; see also Huang, Holcombe, & Pashler, 2004) However, in the present study we found no evidence of any color-dependent interference on position priming, nor of an inhibitory main effect (such as feature-independent IOR), which could have counteracted position priming in Maljkovic and Nakayama (1996), implying that early target selection is immune to these inhibitory effects. Evidence from neurophysiology and neuropsychology suggests that separate anatomical mechanisms account for the priming of different stimulus dimensions. Banissy, Walsh, and Muggleton (2012) observed specific disruptions in color priming when between-trial transcranial magnetic stimulation (TMS) was applied to area V4, but not when stimulating area V5/MT, in healthy participants performing simple visual search tasks. Conversely, stimulating area V5/MT disrupted priming of motion direction but not spatial position (Campana, Cowey, & Walsh, 2006), but stimulating the left (but not the right) frontal eye fields disrupted priming of spatial position, but not priming of motion direction (Campana, Cowey, Casco, Oudsen, & Walsh, 2007). Converging evidence for the modularity of priming effects has come from functional magnetic imaging, in which color and position repetition are related to suppression of the BOLD signal in anatomically distinct areas (Kristjánsson, Vuilleumier, Schwartz, Macaluso, & Driver, 2007; see also Rorden, Kristjánsson, Revill, & Sævarsson, 2011), and from neuropsychology (Kristjánsson, Vuilleumier, Malhotra, Husain, & Driver, 2005). Although functional independence is not a necessary consequence of anatomical dissociation, here we have shown that this is indeed the case during target selection.
Let us note that the color-priming effects reported here go against those of Huang and Pashler (2005), who tied perceptual priming during brief presentation to expected target features, not to repetitions. The present experimental design involved no statistical predictability about upcoming trials. All independent variables (color, position, and target identity) were randomized throughout, excluding any expectation effects. This does not mean that priming of visual search is not affected by expectation (see, e.g., Fecteau, 2007; Fuggetta, Campana, & Casco, 2007; Huang & Pashler, 2005; Kristjánsson, Sigurjónsdóttir, & Driver, 2010), but only that expectation is not necessary for such priming. In many studies of priming, the probability of repetition has been manipulated, making it impossible to dissociate the contributions of recent search history and of the overall statistical properties of a task. The present study shows that priming is strong even without any expectation component. Much evidence has suggested that priming occurs at more than one stage of processing (Ásgeirsson & Kristjánsson, 2011; Campana, Pavan, & Casco, 2008; Lamy, Yashar, & Ruderman, 2010; Lamy, Zivony, & Yashar, 2011; Yashar & Lamy, 2011; see Kristjánsson & Campana, 2010, for a review). A plausible proposal by Lamy et al. (2011, p. 2108) is that feature priming is fundamentally a selection-based phenomenon that is sometimes influenced by response-based effects or, according to the present authors’ own account (Ásgeirsson & Kristjánsson, 2011), by the saliency of an irrelevant feature.
Our results indicate that priming can occur even with very brief stimulus presentation. Even though we cannot make precise inferences about the minimum exposure duration sufficient to produce priming effects, they are obvious following an ~70-ms presentation. A priming pattern is visible in the descriptive data from the briefest above-threshold presentations, suggesting that very brief exposure may suffice for repetition benefits.
To account for the observed repetition-priming patterns, we tentatively propose that, other things being equal, the occurrence of a feature j (e.g., a color or a spatial position) as a feature of a target on trial n increases the pertinence of feature j on trial n + 1 (see Bundesen, 1990, pp. 540ff). Pertinence refers to the π values in Bundesen’s theory of visual attention (TVA). In TVA, π values determine which objects are selected (filtering), whereas perceptual biases (β values) determine how the objects are categorized (pigeonholing). According to the rate and weight equations of TVA (Bundesen, 1990, Eqs. 1 and 2), when the pertinence of feature j increases, the attentional weights of objects containing feature j also increase, which in turn implies an increase in the readiness to see not only feature j, but all sorts of visual features of objects containing feature j (including the letter type of objects with feature j). Note that negative priming (see, e.g., Bundesen & Habekost, 2008, pp. 23f, 129ff) may be explained by a similar hypothesis stating that, other things being equal, the occurrence of j as a feature of a distractor on trial n decreases the pertinence of feature j on trial n + 1.
The validity of the qualitative hypotheses that we have suggested for priming may presuppose a nontrivial selection task and a sufficiently salient feature j, but no particular role of the primed feature (such as the role of being a target-defining feature). Figure 2a shows our data fitted by a simplified TVA model based on these assumptions, such that the rate of processing is most accurate and efficient when both color and position are repeated, lower when only one attribute is repeated, and lowest when both change on a subsequent trial. Further details on state-of-the-art modeling based on TVA may be found in Bundesen and Habekost (in press), but—in line with the qualitative hypotheses stated in the previous paragraph—the fit shown in Fig. 2a is to be considered illustrative rather than a test of the model.
Conclusions
Priming from purely perceptual processes plays a significant and important role in visual selection, increasing the probability of correctly reporting targets, rather than just contributing to faster responses. This is, most likely, due to increases in the probability of encoding targets into visual short-term memory during the brief window of stimulus exposure (see Bundesen, 1990; Bundesen, Habekost, & Kyllingsbæk, 2005). We conclude that postperceptual decision-making cannot explain our results, since (1) our task was a nonspeeded accuracy-based task, which should not involve any artifacts of motor components and/or postperceptual decision-making, and (2) fully randomized presentation contingencies rule out any role of expectations.
Unlike Maljkovic and Nakayama (1996), we found no signs of a modulation of position priming by inhibition of return. Position priming was likely present from more than just the previous trial, but this facilitation was much weaker than that observed from the most recent trial. Most importantly, our results show, for the first time, that the priming effects of feature and position are independent of one another in the identification of briefly presented, postmasked stimuli. This contrasts with much of the literature on repetition in reaction time tasks. However, the conflicting evidence is readily explained by invoking different stages of processing. In the present study, we isolated the target selection stage from postperceptual stages and showed that position and color priming do not interact during selection.
Notes
RMSD is the root-mean squared deviation—that is, the square root of the mean squared error term for each effect.
References
Ásgeirsson, Á. G., & Kristjánsson, Á. (2011). Episodic retrieval and feature facilitation in intertrial priming of visual search. Attention, Perception, & Psychophysics, 73, 1350–1360. doi:10.3758/s13414-011-0119-5
Ásgeirsson, Á., Kyllingsbæk, S., Kristjánsson, Á., & Bundesen, C. (2012). The weight of the visual world is modified by recent experience: Modeling repetition priming in a partial report task. Journal of Vision, 12(9), 931. doi:10.1167/12.9.931
Banissy, M. J., Walsh, V., & Muggleton, N. G. (2012). A disruption of colour priming following continuous theta burst transcranial magnetic stimulation. Cortex, 48, 1359–1361.
Becker, S. I., & Horstmann, G. (2009). A feature-weighting account of priming in conjunction search. Attention, Perception, & Psychophysics, 71, 258–272. doi:10.3758/APP.71.2.258
Brainard, D. H. (1997). The psychophysics toolbox. Spatial Vision, 10, 433–436. doi:10.1163/156856897X00357
Brascamp, J., Blake, R., & Kristjánsson, Á. (2011). Deciding where to attend: Priming of pop-out drives target selection. Journal of Experimental Psychology: Human Perception and Performance, 37, 1700–1707.
Bundesen, C. (1990). A theory of visual attention. Psychological Review, 97, 523–547. doi:10.1037/0033-295X.97.4.523
Bundesen, C., & Habekost, T. (2008). Principles of visual attention: Linking mind and brain. New York, NY: Oxford University Press.
Bundesen, C., & Habekost, T. (in press). Theory of visual attention (TVA). In A. C. Nobre & S. Kastner (Eds.), The Oxford handbook of attention. New York, NY: Oxford University Press.
Bundesen, C., Habekost, T., & Kyllingsbæk, S. (2005). A neural theory of visual attention: Bridging cognition and neurophysiology. Psychological Review, 112, 291–328. doi:10.1037/0033-295X.112.2.291
Bundesen, C., Kyllingsbæk, S., & Larsen, A. (2003). Independent encoding of colors and shapes from two stimuli. Psychonomic Bulletin & Review, 10, 474–479. doi:10.3758/BF03196509
Cambridge Research Systems. (2008). Video eyetracker toolbox: User manual. Rochester, NY: Author.
Campana, G., & Casco, C. (2009). Repetition effects of features and spatial position: Evidence for dissociable mechanisms. Spatial Vision, 22, 325–338.
Campana, G., Cowey, A., Casco, C., Oudsen, I., & Walsh, V. (2007). Left frontal eye field remembers “where” but not “what”. Neuropsychologia, 45, 2340–2345. doi:10.1016/j.neuropsychologia.2007.02.009
Campana, G., Cowey, A., & Walsh, V. (2006). Visual area V5/MT remembers “what” but not “where”. Cerebral Cortex, 16, 1766–1770.
Campana, G., Pavan, A., & Casco, C. (2008). Priming of first-and second-order motion: Mechanisms and neural substrates. Neuropsychologia, 46, 393–398.
Dawkins, M. (1971). Shifts of “attention” in chicks during feeding. Animal Behaviour, 19, 575–582.
Fecteau, J. H. (2007). Priming of pop-out depends upon the current goals of observers. Journal of Vision, 7(6):1, 1–11. doi:10.1167/7.6.1.
Fuggetta, G., Campana, G., & Casco, C. (2007). The principle of good continuation in space and time can guide visual search in absence of priming or contextual cueing. Visual Cognition, 15, 834–853.
Goolsby, B., & Suzuki, S. (2001). Understanding priming of color-singleton search: Roles of attention at encoding and “retrieval”. Perception & Psychophysics, 63, 929–944. doi:10.3758/BF03194513
Hillstrom, A. P. (2000). Repetition effects in visual search. Perception & Psychophysics, 62, 800–817. doi:10.3758/BF03206924
Huang, L., Holcombe, A., & Pashler, H. (2004). Repetition priming in visual search: Episodic retrieval, not feature priming. Memory & Cognition, 32, 12–20. doi:10.3758/BF03195816
Huang, L., & Pashler, H. (2005). Expectation and repetition effects in searching for featural singletons in very brief displays. Perception & Psychophysics, 67, 150–157. doi:10.3758/BF03195018
Kristjánsson, A. (2006a). Rapid learning in attention shifts: A review. Visual Cognition, 13, 324–362. doi:10.1080/13506280544000039
Kristjánsson, Á. (2006b). Simultaneous priming along multiple feature dimensions in a visual search task. Vision Research, 46, 2554–2570.
Kristjánsson, A. (2008). “I know what you did on the last trial”—A selective review of research on priming in visual search. Frontiers in Bioscience, 13, 1171–1181.
Kristjánsson, Á. (2009). Independent and additive repetition priming of motion direction and color in visual search. Psychological Research, 73, 158–166.
Kristjánsson, Á., & Campana, G. (2010). Where perception meets memory: A review of repetition priming in visual search tasks. Attention, Perception, & Psychophysics, 72, 5–18. doi:10.3758/APP.72.1.5
Kristjánsson, Á., Ingvarsdöttir, Á., & Teitsdöttir, U. D. (2008). Object- and feature-based priming in visual search. Psychonomic Bulletin & Review, 15, 378–384. doi:10.3758/PBR.15.2.378
Kristjánsson, Á., Saevarsson, S., & Driver, J. (2013). The boundary conditions of priming of visual search: From passive viewing through task-relevant working memory load. Psychonomic Bulletin & Review, 20, 514–521. doi:10.3758/s13423-013-0375-6
Kristjánsson, Á., Sigurjónsdóttir, Ó., & Driver, J. (2010). Fortune and reversals of fortune in visual search: Reward contingencies for pop-out targets affect search efficiency and target repetition effects. Attention, Perception, & Psychophysics, 72, 1229–1236. doi:10.3858/APP.72.5.1229
Kristjánsson, Á., Vuilleumier, P., Malhotra, P., Husain, M., & Driver, J. (2005). Priming of color and position during visual search in unilateral spatial neglect. Journal of Cognitive Neuroscience, 17, 859–873.
Kristjansson, A., Vuilleumier, P., Schwartz, S., Macaluso, E., & Driver, J. (2007). Neural basis for priming of pop-out during visual search revealed with fMRI. Cerebral Cortex, 17, 1612–1624. doi:10.1093/cercor/bhl072
Kyllingsbæk, S., & Bundesen, C. (2007). Parallel processing in a multifeature whole-report paradigm. Journal of Experimental Psychology: Human Perception and Performance, 33, 64–82. doi:10.1037/0096-1523.33.1.64
Kyllingsbæk, S., Markussen, B., & Bundesen, C. (2012). Testing a Poisson counter model for visual identification of briefly presented, mutually confusable single stimuli in pure accuracy tasks. Journal of Experimental Psychology: Human Perception and Performance, 38, 628–642. doi:10.1037/a0024751
Lamy, D., Yashar, A., & Ruderman, L. (2010). A dual-stage account of inter-trial priming effects. Vision Research, 50, 1396–1401. doi:10.1016/j.visres.2010.01.008
Lamy, D., Zivony, A., & Yashar, A. (2011). The role of search difficulty in intertrial feature priming. Vision Research, 51, 2099–2109.
Luce, R. D. (1963). Detection and recognition. In R. D. Luce, R. R. Bush, & E. Galanter (Eds.), Handbook of mathematical psychology, vol. I (pp. 103–189). New York, NY: Wiley and Sons.
Maljkovic, V., & Nakayama, K. (1994). Priming of pop-out: I. Role of features. Memory & Cognition, 22, 657–672. doi:10.3758/BF03209251
Maljkovic, V., & Nakayama, K. (1996). Priming of pop-out: II. The role of position. Perception & Psychophysics, 58, 977–991. doi:10.3758/BF03206826
Nakayama, K., Maljkovic, V., & Kristjánsson, A. (2004). Short-term memory for the rapid deployment of visual attention. In M. S. Gazzaniga (Ed.), The cognitive neurosciences III (pp. 397–408). Cambridge, MA: MIT Press.
Pelli, D. G. (1997). The VideoToolbox software for visual psychophysics: Transforming numbers into movies. Spatial Vision, 10, 437–442. doi:10.1163/156856897X00366
Pratt, J., & Castel, A. D. (2001). Responding to feature or location: A re-examination of inhibition of return and facilitation of return. Vision Research, 41, 3903–3908.
Rorden, C., Kristjánsson, Á., Revill, K. P., & Sævarsson, S. (2011). Neural correlates of inter-trial priming and role reversal in visual search. Frontiers in Human Neuroscience, 5, 151.
Sigurdardottir, H. M., Kristjánsson, A., & Driver, J. (2008). Repetition streaks increase perceptual sensitivity in visual search of brief displays. Visual Cognition, 16, 643–658. doi:10.1080/13506280701218364
Sperling, G. (1960). The information available in brief visual presentations. Psychological Monographs: General and Applied, 74(11), 1–29.
Treisman, A., Vieira, A., & Hayes, A. (1992). Automaticity and preattentive processing. American Journal of Psychology, 102, 341–362.
Vangkilde, S., Bundesen, C., & Coull, J. T. (2011). Prompt but inefficient: Nicotine differentially modulates discrete components of attention. Psychopharmacology, 4, 667–680. doi:10.1007/s00213-011-2361-x
Vogel, E. K., Woodman, G. F., & Luck, S. J. (2001). Storage of features, conjunctions, and objects in visual working memory. Journal of Experimental Psychology: Human Perception and Performance, 27, 92–114. doi:10.1037/0096-1523.27.1.92
Wang, Z., & Klein, R. M. (2010). Searching for inhibition of return in visual search: A review. Vision Research, 50, 220–228. doi:10.1016/j.visres.2009.11.013
Wolfe, J. M., Friedman-Hill, S. R., & Bilsky, A. B. (1994). Parallel processing of part-whole information in visual search tasks. Perception & Psychophysics, 55, 537–550. doi:10.3758/BF03205311
Yashar, A., & Lamy, D. (2010). Intertrial repetition affects perception: The role of focused attention. Journal of Vision, 10(14):3, 1–8. doi:10.1167/10.14.3
Yashar, A., & Lamy, D. (2011). Refining the dual-stage account of intertrial feature priming: Does motor response or response feature matter? Attention, Perception, & Psychophysics, 73, 2160–2167. doi:10.3758/s13414-011-0182-y
Author note
Á.G.Á. and C.B. were supported by the Center of Excellence Program of the University of Copenhagen. Á.K. was supported by the University of Iceland Research Fund and by RANNIS. The authors thank Heiðrún Hafþórsdóttir and Ómar I. Jóhannesson for assistance in the lab.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Ásgeirsson, Á.G., Kristjánsson, Á. & Bundesen, C. Independent priming of location and color in identification of briefly presented letters. Atten Percept Psychophys 76, 40–48 (2014). https://doi.org/10.3758/s13414-013-0546-6
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13414-013-0546-6