
Abstract
Responding to the location of a target is delayed when the target arises at a position previously occupied by a distractor (ignored-repetition trial), relative to when it occurs at a formerly unoccupied location (control trial) [i.e., the spatial negative priming (SNP) effect]. Speculation has held that recently inhibited (distractor) responses resist future execution (i.e., execution resistance [ER]), and thus cause SNP. Evidence for ER has been reported for identity-based tasks using masked prime distractor events. The purpose of this study was to examine the potential impact of ER on response selection in an SNP task for both nonmasked (traditional) and masked primes. We employed a modified SNP task that included nonmasked and masked target-only and distractor-only visual primes (first trial), along with forced choice and free choice probes (second trial). On free choice trials, a selection bias against the prime-distractor-assigned response was evident (same-hand competition, for both nonmasked and masked primes). This selection avoidance was held to reflect ER operating with inhibited prime distractor responses. Further, inhibitory aftereffect patterns were the same for nonmasked and masked distractor primes, and masking target primes transformed a positive to a negative aftereffect, as predicted by the self-inhibition model of mask function set out by Schlaghecken and Eimer (Psychonomic Bulletin & Review, 11, 463–468, 2004).
Similar content being viewed by others

Aftereffects are commonplace processing outcomes and refer those instances in which the consequences of prior processing come forward to exert an impact on current, related processing (e.g., Berlucchi, 2006). Aftereffects can be positive or negative in direction. Phenomena reflective of the negative aftereffect outcome, such as the inhibition-of-return (IOR), negative-priming, and negative compatibility (NCE) effects, are usually indexed in the form of delayed target responding and have often been attributed to inhibition arising across the perceptual and/or motor domains (e.g., Buckolz, Goldfarb, & Khan, 2004; Rafal, Calabresi, Brennan, & Sciolto, 1989; Rafal, Davies, & Lauder, 2006; Schlaghecken & Eimer, 2004; Schlaghecken, Rowley, Sembi, Simmons, & Whitcomb, 2007). It is important to establish the specific cause (or “functional locus”) of each of these differently labeled aftereffects in order, to ensure that they are distinct inhibitory phenomena, as their separate monikers imply. This causal information can then be used to ascertain what beneficial purpose the processes underlying these aftereffects might serve. More pragmatically, using inhibitory aftereffects as diagnostic tools makes it imperative that we fully understand what processing these aftereffects are reflecting (e.g., Vaughan, Hughes, Jones, Woods, & Tipper, 2006).
The precise causes of the phenomena of inhibitory aftereffects are still being debated (e.g., Berlucchi, 2006; Chao, 2009; Neill, 2007; Sumner, 2007). Accordingly, the first of several motivations for the present research was to establish a more detailed understanding of the processing causing the visual spatial negative priming (SNP) effect.
Defining the SNP effect: its task design and possible cause
In the visual SNP design, events (target, distractor, or both) are presented at centrally positioned locations (foveal region) with trials delivered in pairs—first the “prime” and then the “probe.” Participants are required to manually respond to the location of the target stimulus while ignoring any distractor event, if one is present. When the probe target assumes the location previously occupied by the prime distractor event (i.e., ignored-repetition trial; see Fig. 1, panel 3 vs. panel 6a), reaction times (RTs) and (often) buttonpress error rates are elevated beyond those obtained on control trials, where the probe target arises at a location unused on the prime (e.g., Fitzgeorge & Buckolz, 2008). Although both of these measures are of theoretical importance, the (RT[ignored repetition] > RT[control]) inequality is usually the sole basis for detecting SNP effect presence.

An illustrative sequence for a forced choice (ignored-repetition trial) and a free choice (same hand, distractor response repetition [DRR] vs. control [CO] competition) trial following either a nonmasked or a masked distractor prime. The solid white square represents a to-be-ignored distractor event, and the solid black squares represent to-be-responded-to target events. Locations 1–4 (L1–L4) in the inset are the forced choice location bar markers (1:1 location-to-response mapping), and Locations 5–7 (L5–L7) are the free choice location bar markers (1:2 location-to-response mapping)
Presently, the visual SNP effect is held to evolve during response end processing (Buckolz et al., 2004; Guy, Buckolz, & Khan, 2006; also see Neill, 2007). Presumably, the distractor-occupied prime-trial location automatically sets in motion an activation (A)–inhibition (I)–execution resistance (ER) sequence (A→I→ER) that is applied to its related response (Fitzgeorge & Buckolz, 2008). That is, the prime distractor response is unintentionally activated and then similarly inhibited in order to prevent its unwanted initiation. An automatic outcome of this inhibition is to render the involved response resistant to later execution for a period of time (henceforth, “execution resistance” or ER). This ER feature is responsible for the inhibitory aftereffects observed on IR trials in SNP tasks. On these trials, ER opposes the probe target’s stipulation that the former distractor response be executed. As expected, the probe target usually wins this competition. This victory comes with a time cost that accrues when ER is overridden, a process that delays probe target response-initiation time, which causes the SNP effect.
Alternatively, when ER prevails on ignored-repetition trials, a response selection error is recorded. Ideally, such ER victories over the probe target should be rare, and this is the case. Numerically, the ignored repetition > control differential is small (1%–2%) and often nonsignificant, even when an SNP effect is indicated by the latency measure (e.g., Christie & Klein, 2008; Fitzgeorge & Buckolz, 2008; Gibbons & Rammsayer, 2004; Guy, Buckolz, & Fitzgeorge, 2007; Guy, Buckolz, & Pratt, 2004; Milliken, Tipper, Houghton, & Lupiáñez, 2000). This volatility explains why an ignored repetition > control error rate differential has not been required to index the presence of an SNP effect to date; however, the disregard of this measure has undermined its importance in providing support for the existence of ER. Accordingly, a major objective of the present study was to look for clearer evidence of ER on probe response selection when its manifestation was unopposed by the probe target. To do this, we incorporated free choice probe trials (see below) into the typical forced choice SNP procedure (see Fig. 1 inset; L5–L7 were free choice probe-trial locations).
Objective 1: seek support for the ER property of recently inhibited responses through its impact on response selection on free choice probe trials
With free choice probe trials, the target stimulus delineates a subset of permissible responses (n = 2) between which the participant can “freely” choose (i.e., 1:many location–response mappings). Of particular interest are free choice trials on which an inhibited prime distractor response and a noninhibited control response constitute the allowable probe-trial responses (e.g., in the Fig. 1 inset, when the prime distractor occurs at L2 and the probe target occurs at L5). If ER characterizes a recently inhibited (distractor) response, a clear bias against its selection should emerge on free choice trials, where an ER influence is not opposed/obscured by the probe target.
Free choice probe trials can provide two other instructive results. First, appropriate selection-against data would identify ER as a response-selection determinant on free choice trials. Because free choice episodes are common in everyday life situations (e.g., a driver is confronted with several response options when trying to avoid hitting an obstacle moving into his/her pathway) but are infrequently studied in controlled environments, a delineation of the factors that guide free choice response selection is a worthy goal in and of itself.
Second, free choice probe-trial data will inform us about the functional loci of both inhibitory (distractor prime on ignored-repetition trials) and positive (target prime on target-repeat trials) aftereffects that occur in visual SNP designs. This follows because on free choice trials, the probe target is positioned at a new location (L5–L7, Fig. 1 inset) away from the prime event’s position (i.e., L1–L4, forced choice locations). Thus, aftereffects revealed on free choice trials are more logically attributed to features carried forward by a repeated response (i.e., a response locus) than to those related to a repeated location. For example, on free choice trials that follow a distractor-only prime, on which the distractor and control responses compete for action, a selection bias against the former, along with delayed responding when the distractor response is freely chosen (i.e., a distractor response repetition [DRR] trial), would indicate a response locus for ignored-repetition slowing. Buckolz et al. (2004) tested this possibility in an SNP task. They reported equivalent DRR and ignored-repetition trial latencies, indicative of a response locus for the SNP effect. No evidence of ER was obtained on their free choice trials. However, they used only between-hand free choice trials, so hand preference may have obscured ER manifestation. We eliminated this problem by including within-hand free choice trials.
Furthermore, free choice trials following target-only primes, where the prime target response and a control response compete, will indicate whether individuals select for the just-executed target response (i.e., a free choice selection determinant) and whether this choice coincides with a facilitation effect. Also, we will learn whether the reuse of the target-occupied prime location contributes to the classic target-repetition facilitation observed in past SNP designs (e.g., Buckolz, Avramidis, & Fitzgeorge, 2008; Fitzgeorge & Buckolz, 2008). For example, this would not be indicated if comparable latencies were obtained for free choice target response repetition (TRR trials) and for trials on which both target location and response are reused on the probe (target-repeat) trial. Rather, it would signify that response reuse contributes to target-repeat facilitation.
In sum, our within-hand free choice trials will provide new data bearing on (a) identifying the precise cause of the SNP effect, through possible support for the selection-against hypothesis (i.e., ER); (b) identifying response-selection determinants in free choice situations; (c) verifying or establishing the loci responsible for inhibitory (ignored-repetition) and positive (target-repeat) aftereffects, respectively, in SNP tasks; and (d) determining whether freely selecting responses alters their aftereffect characteristics relative to when the use of these outputs is required.
The NCE: existing support for ER
Results from NCE studies have provided support for the existence of ER. To illustrate, the basic NCE task delivers an uninformative, to-be-ignored cue (e.g., double arrows, << or >>) of brief duration (~16 ms) at a central fixation point (i.e., a prime trial), with its offset immediately followed by a mask (e.g., superimposed cue arrows) that lasts about 100 ms. When the mask terminates, it is followed after various delays by the probe-trial target (i.e., << or >>), indicating the need for a left- or right-hand buttonpress response, respectively. On these forced choice trials, reaction times are faster when the prime and probe responses match (compatible trial; i.e., a positive compatibility effect) when the prime–probe delay is short; however, this reverses when the delay is extended sufficiently (i.e., the NCE). When free choice trials are also included on the probe trials, individuals exhibit a small but significant bias against selecting the distractor/compatible response (Klapp & Haas, 2005; Klapp & Hinkley, 2002; Schlaghecken & Eimer, 2004; Schlaghecken, Klapp, & Maylor, 2009), reflecting the presence of ER and its involvement in NCE production.
The foregoing brief description reveals two important procedural differences between the NCE and SNP paradigms that seriously puts in question the interchangeability of the aftereffects produced by these two tasks. Specifically, unlike the SNP paradigm, the salient target feature for the NCE task is its identity rather than its location, and the prime distractor has been masked when an NCE occurs. First, identity-based and location-based variants of the visual negative-priming task have not always exhibited similar inhibitory aftereffects in response to the same experimental manipulation (e.g., Buckolz, Boulougouris, & Khan, 2002; Moore, 1994; see Chao, 2011, for additional examples). Second, the concern with the masking component of the NCE procedure in part is that the NCE itself—and so the ER evidence—occurs only when the distractor prime has been appropriately masked. Clearly, unlike in the SNP phenomenon, the NCE is a mask-produced entity. It follows too that the ER data seen with masked primes may not be evident with the traditional, nonmasked primes of the SNP procedure (Objective 1).
Given this possibility, and the lack of published data on the matter, we undertook a second objective—namely, the examination of aftereffects for masked distractor-only and target-only primes in an SNP task.
Objective 2: to examine the inhibitory and positive aftereffects produced by masked distractor and target primes in an SNP task
The self-inhibition account of mask function in NCE tasks
To illustrate the benefits to be gained via Objective 2, some speculation is required as to how mask processing works. Although other accounts exist (see Sumner, 2007), the self-inhibition model of mask behavior advanced by Schlaghecken and colleagues is the alternative preferred here (see, e.g., Schlaghecken & Eimer, 2004; Schlaghecken et al., 2007, 2009). According to this model, briefly exposed distractor events that lack cognitive awareness nevertheless automatically activate their related responses. The timely delivery of an appropriate mask eliminates the sensory representation of the prime-trial distractor event, which can have two consequences. One is to maintain the phenomenal invisibility of the brief prime distractor cue. The other is to trigger a reflexive self-inhibition of a distractor-related response (if it is sufficiently activated), because it has now lost its perceptual support, and so its legitimacy. The time needed to overcome this self-inhibition (perhaps mediated by ER) causes the NCE.
Possible benefits of examining masked prime events
The first benefit is that the materialization of inhibitory aftereffects following successfully masked prime distractors would indicate both that the “location” property of phenomenally invisible events is identified, which then serves to activate the location’s assigned response (later to be inhibited), and that this identification occurs automatically. Regarding the latter point, the processing of the prime distractor event and its related A→I→ER response processing following visible prime distractors has been presumed to occur automatically. This is because manual responses linked to these events have exhibited electrophysiological evidence of enhanced activation (e.g., De Jong, Liang, & Lauber, 1994; Eriksen, Coles, Morris, & O’Hara, 1985; Valle-Inclan & Redondo, 1998) and because this activation presumably occurs without intention. Evidence of the A→I→ER sequence following masked distractor events would constitute more convincing evidence of its automated property (McCormick, 1997; Mulckhuyse & Theeuwes, 2010; Sumner, 2007), because events that do not reach the level of cognitive awareness cannot be a source of volitional processing.
Second, we would be able, for the first time, to contrast the inhibitory aftereffect data for nonmasked and masked distractor primes in an SNP design. Firm predictions in this regard are not warranted, other than to say that inhibitory aftereffects for masked distractors are predicted by the self-inhibition model (Schlaghecken et al., 2007), as well as evidence of ER.
Finally, the self-inhibition model also predicts that successfully masked prime targets will produce inhibitory rather than (nonmasked) positive aftereffects, comparable to those obtained for masked distractor primes, including selection against on free choice trials (see Objective 1). This would occur because the sensory representations of both events would undergo removal, and so would be processed similarly. Obtaining such target results would support the self-inhibition model and show that the mask is in control when introduced into the SNP design, even though an SNP effect arises without the mask (unlike in the NCE task).
Method
A total of 18 right-hand-dominant undergraduate students, ranging in age from 18–30 years and with normal or corrected-to-normal vision, participated in this experiment.
Apparatus
The input display was presented in a dimly lit room on a 47.5-cm computer screen, situated on a tabletop 73.5 cm above the floor, located 190 cm directly in front of the participant (see Fig. 1). The display consisted of two rows of bar markers centered horizontally on the screen and separated vertically from one another by 2.2 cm (0.6º), which specified the possible target (T) or distractor (D) locations. The to-be-responded-to target and the to-be-ignored distractor rectangles were the same size (0.9 cm wide and 1.9 cm high) but differed in color, the target being green ([G:250] + [R: 0] + [B:0]) and the distractor being red ([G: 0] + [R:250] + [B:0]).
The bottom row contained four bar markers (denoted L1–L4 in the Fig. 1 inset, from left to right) that served as the forced choice locations, with each bar marker being assigned a single spatially compatible buttonpress response. The middle and index finger of the left and right hands rested on keyboard buttons D, V, L, and M, respectively, which were assigned to their spatially compatible bar marker locations (L1, L2, L4, L3). The bar markers appeared white in color against the black background and measured 0.9 cm in width. L1 was separated from L2, and L3 from L4, by a distance of 0.8 cm, whereas L2 was separated from L3 by 1.7 cm. The horizontal distance between L1 and L4, center to center, measured 5.9 cm, a horizontal visual angle of about 2º.
The top row contained three yellow location bar markers (denoted L5–L7; see Fig. 1 inset). Bar marker L5 was located midway between bar markers L1 and L2, with bar makers L6 and L7 being positioned midway between L2 and L3, and L3 and L4, respectively. The horizontal distance of the display from L5 to L7, center to center, measured 4.4 cm, yielding a horizontal visual angle of about 1.5º. Locations L5, L6, and L7 served as the free choice locations and were mapped onto both of keyboard buttons D and V, V and M, and M and L, respectively. When the probe target stimulus appeared at a free choice location, participants were “free” to select either of the two responses mapped onto that location. Bear in mind that having the probe target appear at a location other than the prime distractor position does not remove the SNP effect (e.g., Guy et al., 2006) and so should not do so with our free choice trials (Buckolz et al., 2004).
When a probe target stimulus showed up at L5 or L7 following a prime-trial distractor event, the competing finger responses were on the same hand and represented either a prime distractor repeat response (DRR) or a control (CO) response, or, if the distractor appeared on the other side, a pair of control outputs. In contrast, a target stimulus arising at L6 generated a competition between fingers on different hands, again consisting of either DRR versus CO or CO versus CO response types. The important point is that the same-hand DRR versus CO competitions allowed us to examine the inhibitory aftereffect impact on free choice response selection when hand dominance was not a selection determinant, a problem that has arisen in the past in a location SNP task (Buckolz et al., 2004). Alternatively, the between-hand CO versus CO condition allowed an assessment of hand dominance on response selection without the involvement of aftereffects. These data were only of peripheral interest (Appendix A).
When a probe target stimulus showed up at a free choice location (L5, L6, or L7) following a prime-trial target stimulus, the competing finger responses represented either a previously executed target response (TRR) or a control (CO) response. Again, a probe-trial target stimulus appearing on L5 or L7 generated competitions on the same hand (TRR vs. CO or CO vs. CO), and a target stimulus arising at L6 generated a different-hand competition (TRR vs. CO or CO vs. CO).
The term “free choice” denotes those occasions on which the probe target stimulus permitted the use of more than one allowable response (e.g., Schlaghecken & Eimer, 2004). It is not intended to imply limitless response alternatives, free selection among all possible experimental responses, nor choice without constraints.
Also appearing on the computer screen, below the white forced choice response location bar markers, was a schematic or stick figure (overhead view) of a participant. The lines representing forearms ended in Vs, representing the third digit and index finger on both hands. These “digits” extended within 1 cm of the forced choice response bar markers to which they were, respectively, assigned. The stick figure (Fig. 1 inset) was included to ensure that each participant could clearly distinguish among the bar marker positions and has been used in prior work (e.g., Guy & Buckolz, 2007; Guy et al., 2006).
Procedures
Trials were presented in pairs, first the prime and then the probe trial. An illustrative sequence of a trial pair is presented in Fig. 1, along with event timing. All trials began with a warning tone (100 ms) whose offset was followed by the display configuration, which remained on the screen for the entire prime–probe trial sequence (panel 2). The prime-trial event (panel 3), either a single distractor (D) or a single target (T), appeared 400 ms after the configuration onset and remained on the screen for a duration of 16 ms (Eimer & Schlaghecken, 2002). Prime events occurred only at the forced choice locations (L1–L4) and were presented equally often and randomly, with the restriction that they appeared with equal frequency at all four of the forced choice bar markers. Participants were required to ignore distractor-occupied locations and to respond with the appropriate buttonpress response to target positions whenever a target was detected, and this held irrespective of whether the primes were masked or not.
When used, the prime mask was delivered coincident with the offset of the prime-trial event and covered all bar marker locations remaining on the screen for 100 ms (panel 4b). The mask itself was a solid yellow rectangle ([R: 255] + [G: 255] + [B: 0]) that had shown success in maintaining the phenomenal invisibility of our prime distractor and target events in a pilot study.Footnote 1 The presentation of masked and nonmasked prime trials was blocked and experienced in a counterbalanced order.
A total of 700 ms following the offset of the mask, or 800 ms following the offset of the prime-trial event (no mask), the probe-trial display was delivered. The single probe-trial event (either target [green rectangle, 94%] or distractor [red rectangle, 6%]; see panel 6a [forced choice] and panel 6b [free choice, same hand]) lasted 100 ms and appeared randomly at one of the seven locations. A block included a mixture of forced choice (n = 512) and free choice (n = 384) probe trials, as well as 56 catch trials (total of 952 trials per block). Participants completed a block in two visits (45 min each) separated by at least 1 day, and undertook one trial block each for the masked and no-mask primes (counterbalanced order). Thus, participants completed a total of 1,904 trials overall.
The response types (ignored repetition [IR], DRR, target repetition [TR], TRR, and CO) and their respective frequencies for the forced choice and free choice trial types, along with a further breakdown for within same-hand and between different-hand competitions, are presented in Appendix A. Recall also that on free choice trials, competitions could be between DRR and CO response types, between TRR and CO response types, or between two CO response types. Trial breakdowns are presented for these subconditions as well.
Following a probe distractor or a correct probe target response, the computer screen became blank, signifying the end of the trial pair and concurrently initiating the 1,500-ms intertrial interval. The termination of this interval coincided with the presentation of the warning tone and the beginning of another prime–probe pair sequence.
Participants were automatically offered rests after every 40 trial pairs. A trial series was restarted when the participant pressed the space bar. Participants could also press the escape button to institute a break at any time.
Participants received the following instructions: (1) to respond as quickly as possible to any target stimulus (green rectangle) while ignoring any distractor (red rectangle) that might be present and to keep buttonpress errors to a minimum; (2) to complete 10 practice trial pairs and ask task requirement questions prior to starting the experimental session; and (3) to avoid preplanning and developing strategies for free choice responses.
Data analyses
Free choice response selection data
Chi-squared values were calculated using 2 × 2 contingency tables formed by the Mask Type (no mask vs. mask) and Response Type factors for same-hand finger pairings. These computations were done separately following distractor-only and target-only primes, so, accordingly, the components of the Response Type factor for these two analyses differed. For the chi-squared analyses following a distractor prime, the Response Type factor consisted of the prime distractor location’s assigned response (DRR) versus a control (CO) response (see, e.g., Fig. 1, panels 3 and 6b; DR = left index, CO = left third digit). Following a target prime, the Response Type factor consisted of a repeated target response (TRR) and a control (CO) response (e.g., in Fig. 1, the event in panel 6b is the target event, hence the free choice trial contest in that panel would consist of a target response [left index] and a CO response [left third digit]). Thus, in the case of free choice trials, both the target object (i.e., the green rectangle) and the primed target response are reinvolved on TRR trials.
For different-hand finger pairings, unless otherwise indicated later, chi-squared calculations were done using Response Type and Hand Preference as the main factors, and were computed separately following target-only and distractor-only primes. Again, then, the components of the response factor varied, consisting of DRR versus CO or of a pair of CO responses subsequent to distractor-only primes, or of TRR versus CO or a pair of CO responses following a target-only prime.
Reaction time data
Within-subject mean probe reaction times were submitted to ANOVA calculations with Aftereffect, Mask Type (no mask vs. mask), and Choice Condition (forced vs. free choice [same hand]) serving as the main factors. Separate ANOVAs were computed following distractor-only and target-only primes. The Aftereffect factor contrasted ignored-repetition with control trial reactions following distractor-only primes and target repetitions with control latencies subsequent to target-only primes. A parallel ANOVA was carried out for between-hand finger competitions, with the exception that the choice condition consisted of the left-hand versus right-hand index finger responses.
Buttonpress error data
ANOVA calculations for the buttonpress error rates on forced choice trials had Aftereffect and Mask Type as main factors.
RTs of less than 100 ms (anticipations) or greater than 1,200 ms (insufficient vigilance) [less than 1% of all trials administered], along with keyboard buttonpress errors, were excluded from RT analyses, as were probe RTs following responses to masked prime events. Finally, examination of the selection-against hypothesis was restricted to same-hand finger response competitions in order to avoid hand dominance being a confounding selection determinant on free choice trials (see Buckolz et al., 2004). For consistency, the nonmasked versus masked prime (latency-indexed) aftereffect comparisons with free choice trials discussed in the main text are also based only on same-hand competitions. Between-hand data are nonetheless presented with the same-hand results for ease of comparison, and for the interested reader, analyses associated with these results, along with a brief discussion, are presented in the Appendix B.
Prime-trial results
When no mask was used, participants consistently responded to the appearance of the prime target stimulus and consistently withheld responding to prime distractor events. Distractor primes elicited a response on less that 1% of these trials, and target primes generated a manual response that was accurate 97% of the time. In contrast, when the prime events were masked, the presentation of the single target stimulus rarely led to an overt response (2.85%); this despite the fact that the salience of the target object, as a to-be-responded-to event, was reinforced on each probe trial. Also, participants’ verbal reports at the end of the study generally indicated that they were unaware of either distractor or target events appearing on the prime trial. The data for one participant were deleted due to excessive response rates to masked prime targets.
These results converge with those of our pilot study (see note 1) to show that the phenomenal invisibility of our prime events was maintained by the masking procedure.
Probe-trial results: distractor primes
Response selection on free choice probe trials
Free choice response selection data are presented in Table 1 and Fig. 2 (as percentages deviant from chance). The detailed different-hand competition results are presented in Appendix A.

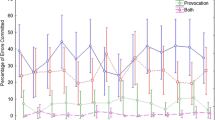
Free choice response selection bias (%): Frequency of avoiding (selection against) versus favoring (selection for) the response assigned to the presented prime event (distractor prime or target prime), following both nonmasked and masked primes. *p < .05
Same-hand competitions: DRR versus CO
A significant chi-squared value was obtained, χ 2(1, n = 2,176) = 5.36, p < .05, when the competing finger responses were on the same hand. Follow-up tests indicated that individuals chose the control response significantly more often than the primed distractor response for both the nonmasked, χ 2(1, n = 1,088) = 63.09, p < .05, and masked, χ 2(1, n = 1,088) = 124.17, p < .05, primes. This imbalance was reliably larger (p < .05) following a masked distractor prime location (.67/.33) than when no mask was delivered (.62/.38); however, in quantitative terms, this difference was small.
Latency data
The cell means for this ANOVA are found in Table 2. Aftereffect scores are presented in Fig. 3.

Latency-indexed aftereffect values (in milliseconds): An illustration of the inhibitory and positive aftereffects for forced choice and free choice trials following masked or nonmasked primes (different hands combined vs. same hand). Aftereffects equal the latency differences between ignored repetition, distractor-response repetition, target repetition, and target-response repetition trials and their appropriate control reaction times. *p < .05
Unqualified significant main effects showed that probe-trial reactions were significantly faster following a masked prime (390 vs. 438 ms), perhaps owing to an alerting impact associated with the mask presentation, F(1, 16) = 42.15, MSE = 1,850, p < .01, and were reliably slower for free choice (422 ms) than for forced choice (407 ms) trials, F(1, 16) = 5.95, MSE = 1,196, p < .05. Notably, aftereffect did not interact with the choice condition, F < 1, but it did interact with mask type, F(1, 16) = 10.80, MSE = 568, p < .01. Follow up post-hoc tests revealed the presence of a significant inhibitory aftereffect for masked prime events (35 ms); however, its size was significantly reduced relative to the effect produced when the mask was absent (61 ms).
No other significant effects were observed. The aftereffect sizes were comparable for forced choice (48 ms) and free choice (47 ms) trials.
Buttonpress errors
Buttonpress error rates were numerically larger for ignored-repetition than for control trials on forced choice trials. This was evident for both nonmasked (2.04% vs. 1.58%) and masked (2.22% vs. 1.66%) distractor primes; however, ANOVA calculations showed that the trial type main effect only approached statistical significance, F(1, 16) = 3.78, MSE = 1.17, p = .070.
Discussion: distractor primes
Same-hand competitions
Nonmasked distractor primes
Free choice trials following visible primes supported an output locus for the traditional SNP task in two ways. First, latencies following visible primes for trials on which the participants freely chose the prime distractor response (i.e., DRR trials) significantly exceeded the latencies obtained for self-selected control responses. This latency difference (RT[DRR] > RT[CO]) was equivalent to the difference seen on forced choice trials (RT[ignored repetition] > RT[control]; i.e., the SNP effect). Collectively, these findings make it clear that the use of the former distractor response on the probe was sufficient to produce the SNP effect (i.e., response locus), without including reuse of the prime distractor location (Buckolz et al., 2004). Second, participants exhibited a significant bias against selecting the prime distractor response on free choice trials (67%) when it competed with a control (noninhibited) response. This selection-against bias presumably reflects the influence of an ER property automatically imposed on recently inhibited responses that acts to prevent a response’s later use. Seemingly, it takes time to successfully counteract ER on ignored-repetition trials, which then produces the SNP effect.
The clear presence and operation of ER on free choice trials was barely evident on buttonpress error rates (ignored-repetition > control), for which ER is also held to be the cause. This muted manifestation is typical (see, e.g., Buckolz et al., 2008; Fitzgeorge & Buckolz, 2008); it accounts for the lack of ER evidence in the past and is attributable to the fact that, unlike on free choice trials, the impact of ER on response selection is opposed and dominated on ignored-repetition forced choice trials by the dictates of the probe target.
On a different note, it bears highlighting that although the inhibitory aftereffects caused by ER in SNP tasks can be set aside in some circumstances (e.g., Buckolz et al., 2002; Fitzgeorge & Buckolz, 2008), this does not occur with self-selected distractor response use. This failed modulation has also been reported for identity-based NCE tasks (e.g., Schlaghecken & Eimer, 2004). From an optimal design perspective, this modulation absence seems maladaptive. Ideally, freely choosing a response for use should include the removal of any associated inhibitory aftereffects.
Masked distractor primes
It also bears noting that the appearance of inhibitory aftereffects at masked prime distractor locations was predicted by the self-inhibition model of mask function (Schlaghecken et al., 2007); however, more important are the implications of this finding, which are twofold and related. First, we see that subliminal processing can extend beyond stimulus identification (NCE tasks) to include location discrimination, since this differentiation is a prerequisite for the production of inhibitory aftereffects at masked distractor locations. Actually, subliminal location discrimination has already been indicated for (masked) peripheral distractor presentations in IOR-type tasks (see Mulckhuyse & Theeuwes, 2010, for a thorough review). What we add here is that subliminal discrimination occurs not only when locations are easily distinguished from each other (i.e., peripheral), but also when they are less distinct (i.e., central).
Second, the inhibitory aftereffects found here are the first convincing evidence that the prime distractor and its consequent response processing (i.e., A→I→ER) can occur automatically in location-based tasks. Earlier speculation claimed that automatic distractor processing following visible primes in SNP tasks was based on the fact that it had likely occurred unintentionally. It was possible, though, that the visible distractors here may have contributed a volitional component to the prime-distractor-related processing. This drawback is absent with successfully masked distractors; that is, events about which individuals are unaware cannot induce volitional processing (McCormick, 1997; Mulckhuyse & Theeuwes, 2010; Sumner, 2007).
The automated nature of location processing is further indicated by the fact that it takes place even when location discrimination is not needed to perform the task (e.g., on IOR tasks [Mulckhuyse & Theeuwes, 2010] or on Simon tasks [Hommel, 1993]). The utility of such information was illustrated by Buckolz, Fitzgeorge, and Knowles (2011), who found no inhibitory aftereffects in a one-response task using central locations. They claimed that this absence did not result because location discrimination had not occurred; they noted that it took place automatically, in spite of task demands that did not require it. Based on this result, these authors suggested that IOR and SNP have distinct causes.
More generally, our masked prime distractor data are in line with NCE reports (e.g., Schlaghecken et al., 2007). Collectively, they show that events that do not reach the level of conscious awareness can selectively activate their related responses in both location-based and identity-based tasks, possibly by setting the A→I→ER sequence into motion, which culminates in inhibitory aftereffects. These results oppose the view that response inhibition can only occur with visible events (e.g., Allport, Tipper, & Chmiel, 1985; Neill, Valdes, & Terry, 1995).
Finally, turning to comparisons, the inhibitory aftereffect patterns produced with nonmasked and masked distractor primes were highly similar (see Table 2 and Fig. 3). The exception was that the size of the latency-based aftereffects was greater in the former instance. This held for forced choice (62 vs. 34 ms) and free choice (65 vs. 33 ms) trials. The reason for this size difference is unclear. It may signal the involvement of different inhibitory systems for nonmasked and masked distractors. Alternatively, it may reflect that the latter produce lesser levels of response activation, ultimately resulting in reduced ER strength, faster override time, and smaller inhibitory aftereffect magnitudes.
Although it is helpful to learn that inhibitory aftereffects produced by nonmasked and masked distractor primes are largely predictive of one another for location-based tasks, the reason for this similarity is speculative, and so will be considered in the General Discussion. For now, a clear benefit of the aftereffect similarities is that they reinforce the conclusions outlined above associated with nonmasked distractor primes, which are particularly helpful in supporting the existence of ER and the SNP response locus.
Probe-trial results: target primes
Response selection on free choice trials
Same-hand competitions: TRR versus CO
A significant chi-squared value was obtained, χ 2(1, n=2,176) = 171.27, p < .05, reflecting an interactive effect in which individuals selected the prime target response significantly more often than a competing control response when the prime target was not masked, χ 2(1, n=1,088)=63.09, p < .05 (.62 vs. .38). This selection bias was reversed when the prime target was masked, χ 2(1, n=1,088)=111.31, p < .05 (.34 vs. .66; see Table 3 and Fig. 2).
Latency data
A significant aftereffect main effect was qualified by its reliable interaction with mask type, F(1, 16)=19.34, MSE=743, p < .01. Follow-up Newman–Keuls tests (p < .05) revealed that when no mask was used, target-repeat reactions were faster than those for control trials (−61 ms); however, this ordinal relationship was reversed when the prime target was masked (+36 ms). No other significant effects were obtained. Hence, aftereffect sizes were the same for forced and free choice trials within a mask condition. See Table 4 for the detailed results.
Buttonpress error rates
The ANOVA produced no significant effects.
Discussion: target primes
Same-hand competitions
Nonmasked target primes
Under the forced choice condition, nonmasked target primes produced a significant, latency-indexed positive aftereffect on target-repeat as opposed to control trials (Fig. 3; see, e.g., Baylis, Tipper, & Houghton, 1997; Buckolz et al., 2008; Fitzgeorge & Buckolz, 2008; Guy et al., 2004, 2006, 2007; Neill, Terry, & Valdes, 1994; but see Milliken et al., 2000). The target object, location, and response were all reused on a target-repeat trial, while only the target object was reused on a control trial. Thus, the positive aftereffect observed indicates that reusing a former target’s location and/or response contributes to a facilitation effect.
On free choice trials, individuals exhibited a significant tendency to select the prime target response when it competed with a control response, which resulted in a reliable positive aftereffect. The size of this positive aftereffect equaled that produced on forced choice target-repeat trials. Two conclusions follow. First, response reexecution, but not target location reuse, contributes to positive aftereffects. When only the prime target response is reexecuted, a positive aftereffect of equal size arises whether the prime target position is reoccupied or not. Like distractor responses, target response self-selection does not alter the aftereffect influence produced when the response is forced. Second, recent response executions are a free choice selection determinant.
Masked target primes
Most importantly, masked target primes produced inhibitory rather than positive aftereffects that were identical to those seen for masked distractor locations (i.e., target responses were selected against, and their use produced delayed reactions; see Figs. 2 and 3, respectively, for the forced and free choice trials). This transformation is predicted by, and so supports, the self-inhibition model of mask operation set out by Schlaghecken et al. (2007). The self-inhibition model holds that masked processing aftereffects should be inhibitory in direction and saliency independent, since both masked target and distractor events would have their sensory representations removed. This would cause their related responses to be self-inhibited. If this is so, the target aftereffect transformation also shows that self-inhibition takes over when masked primes are introduced into an SNP procedure that already produces inhibitory aftereffects.
General discussion
The cause of the visual SNP effect
The processing giving rise to the SNP effect is now clearer. Visible prime distractors, whether they compete with a target or not (Buckolz et al., 2008), undergo automatic processing that includes the activation and subsequent inhibition of their related responses (Buckolz et al., 2008). Owing to the latter effect, these responses then become resistant to future execution (i.e., an ER feature). The use of the prime distractor response, dictated by the probe target on ignored-repetition trials, requires a time-consuming overriding of ER that extends ignored-repetition RTs beyond those seen for control trials, resulting in the SNP effect. Occasions where ER has successfully opposed the probe target would also explain greater buttonpress error rates on ignored-repetition than on control trials when this occurs (e.g., Fitzgeorge & Buckolz, 2008).
The foregoing processing scenario was partially indicated by earlier results, replicated here, that showed that the SNP effect has a response locus. Specifically, it was reported that the imposed (Guy et al., 2006) or self-selected (present free choice trials) use of the former prime distractor response produced equivalent inhibitory aftereffect sizes whether the prime distractor location was reused (ignored-repetition trials) or not. The present study advances our insight by providing the first convincing evidence (a) that distractor processing is automated—that is, the inhibitory aftereffects are the same for nonmasked and masked distractor primes—and (b) that ER exists. In the latter regard, when a previous distractor and a control response competed for action control on free choice (within-hand) trials, individuals showed a significant bias against choosing the former response, presumably reflecting the repelling force of ER.
The response-end SNP explanation sketched above provides against other causal views of visual SNP—namely, that it is caused by (a) location inhibition, (b) event mismatches at the same location on consecutive (ignored-repetition) trials (Park & Kanwisher, 1994; but see Mayr, Hauke, & Buchner, 2009, for auditory SNP exception), or (c) orientation inhibition arising during the perceptual processing of centrally placed distractors (i.e., the same cause as IOR; e.g., Christie & Klein, 2008; Milliken et al., 2000). The critical result pointing to these contentions is that comparable inhibitory aftereffects appear when a former distractor response is used, irrespective of whether the probe target appears at the distractor location or a new one (either many:1 or 1:many location–response mappings; DRR trials). With DRR trials, prime distractors and probe targets appear at different positions, obviating mismatches at a fixed location as well as participants’ having to reorient to the distractor location, thereby removing any orientation inhibition contribution to DRR delays. Also, the observation that distractor locations themselves are not inhibited in a way that contributes to the SNP effect has also been shown for IOR tasks (Fitzgeorge & Buckolz, 2009; Klein, Christie, & Morris, 2005), so this may be a common feature of location processing across tasks.
Finally, although ER evidence had already been reported for identity-based NCE tasks (e.g., Klapp & Hinkley, 2002; Schlaghecken & Eimer, 2004), our study was its first demonstration for unmasked (visible) and masked distractors in a location-based task. Beyond explanation of the SNP phenomenon, our selection-against data showed that the inhibitory aftereffects match for the NCE and SNP tasks. This similarity indicates that ER operates somewhat broadly, with the A→I→ER processing sequence for distractor responses being initiated whether target identity or location is salient. Harvey (1980) also implied a similar mechanism for categorical inhibitory effects. Continuing with the question of breadth, we consider below whether ER involvement extends beyond buttonpress responses.
Inhibition, ER, and aiming responses
The question is whether ER operates more broadly, to include rapid aiming responses. The answer begins with the realization that in this study we have shown that ER can be tied to inhibited limbs, since our buttonpress finger responses lacked an appreciable trajectory (the flexion action might have also been inhibited, but this is unlikely, since it was common to all limbs). Theoretically, ER might operate on the limb of an inhibited aiming response urge; more uncertain, however, is whether ER would be a characteristic of the limb’s trajectory phase. That is, would the aiming trajectory previously used to travel to a location change (ER-induced aversion) when this location has just been associated with a distractor?
Reports dealing with this question are virtually nonexistent. What has been established, however, is that the trajectory component of distractor-induced aiming responses is inhibited, both for oculomotor (see Van der Stigchel, 2010, for a review) and manual (Welsh, 2002) aiming responses. This inhibition opens up the possibility that ER is also attached to trajectories. The problem with this prior work, however, regarding our interest in ER, is that such trajectory inhibition has been tested almost exclusively using a different response (trajectory) from the one presumed to have been inhibited. In this instance, the path to a new location deviates away from the distractor’s position, provided that inhibition of the distractor-induced aiming response has been completed before target response initiation occurs. This finding, however, is deemed to result from a response programming interaction between target and distractor trajectory urges. The suppression of the latter causes the influence of the target urge to be exaggerated, producing a net trajectory that looks to take the movement away from the distractor location. This explanation takes no account of ER. This neglect may be correct, because target movements sometimes veer toward the distractor location, presumably when distractor-generated aiming urges are not fully inhibited at the time of target response initiation. ER could not explain this finding.
In contrast to the above, Welsh (2002, Study 2) presented a single prime distractor event and had individuals later move either to this or to a different location. Evidence of trajectory inhibition was seen in the latter but not in the former case. Hence, there was no evidence of ER causing a trajectory change when moving to the former distractor position. Welsh did, however, find a significant increase in RTs in this case. The added delay may have reflected the time needed to offset an ER-caused resistance to use the distractor (inhibited) trajectory and the limb associated with this urge.
In any event, a more definitive test of the possibility that ER is associated with the inhibited trajectories of aiming responses would be to see whether distractor-induced response urges (on distractor-only prime trials) were selected against on free choice trials, where a single limb made all responses. It would also be of interest to see whether the self-selected nondistractor response exhibited trajectory inhibition and whether the RT and trajectory data obtained by Welsh (2002, Study 2) for forced movements to the prior distractor location could be replicated.
What is the utility of ER to response processing?
Understandably, explanations have been advanced to explain how the processing held to underlie various inhibitory aftereffect phenomena benefits cognition (i.e., do they represent “intelligent design”?). A common theme is that this underlying processing involves protection of one kind or another. For example, the consensus among IOR researchers is that the biphasic RT pattern (i.e., early facilitation, later interference) produced by targets appearing at a previously cued location reflects two benefits (see Mulckhuyse & Theeuwes, 2010); one is an automatic orientation of attention toward a peripheral stimulation, aimed at inspecting this position for potential danger. The other is improved search efficiency. Efficiency improves because orientation inhibition becomes associated with formerly cued locations that resist reorientations (i.e., possibly an ER effect tied to orienting response urges; see Rafal et al., 2006). Hence, already inspected locations containing distractor information are less likely to be revisited. Importantly, these benefits apply only to static environments where the relevancy status of locations/responses remains constant (Klein, 2000).
The cognitive protection benefits of response inhibition, and now of ER, have been less well considered for SNP tasks. One thought is that the selection-against impact of ER should protect against the unwanted use of the prime distractor response on the probe trial. This protection would be mildly pervasive, operating on control trials when the probe distractor arose at its prime location (distractor-repeat trial) or a new position, or when it was absent altogether (assuming that all nontarget responses compete for execution). Another thought, based on the fact that the SNP process is absent (i.e., turned off) when the probe trial predictably lacks a distractor (e.g., Buckolz et al., 2002; Fitzgeorge & Buckolz, 2008; see Moore, 1994), is that negative priming during processing assists by guarding against distractor response execution. Unless additional assumptions are made, this benefit would be highly restricted, applying only to distractor-repeat control trials. Unfortunately, neither of these suggestions has been experimentally verified. To do this, we would need to see prime and/or probe distractor responses underrepresented in probe-trial error rates.
Finally, even if ER’s limited opportunity to reduce probe-trial response errors, noted above, is ultimately verified, the utility of this benefit is nonetheless restricted because it is only functional in static environments. Consequently, ER’s protective utility declines, while its interference impact increases, as the opportunity for relevancy reversals increases (i.e., ignored-repetition trials). In that case, we have some “good news, bad news” scenarios. The good news is that the unhelpful ER consequences in dynamic environments can be muted somewhat, because ER effects can be disengaged when ignored-repetition trials are sufficiently predictable (e.g., Fitzgeorge & Buckolz, 2008). The bad news is that this disengagement may not occur, such as when the probe contains a distractor event (Fitzgeorge & Buckolz, 2008). The good news is that, to the extent that free choice opportunities exist, individuals show an inclination to avoid using just-inhibited responses, and so avoid their interference effects. The bad news is that when an inhibited response is selected, ER’s inhibitory aftereffects remain (Schlaghecken & Eimer, 2004). This persistence is maladaptive and puzzling. Ideally, the certainty that an ER-affected response is about to be used should result in removal of the ER aftereffect. Overall, ER’s utility seems limited. Its possible protection against response errors is restricted and still needs to be verified experimentally, while its interference with target reactions is a sizeable detriment.
Nonmasked versus masked distractor aftereffects in location-based tasks
The unexpected comparable inhibitory aftereffect patterns produced by nonmasked and masked distractor primes possibly arose because automatic response activations are governed by reflexive self-inhibition. This holds for visible and invisible primes (present data). Despite the facts that self-inhibition would likely be mediated by different neural pathways (Brown, Almeida, Fitzgeorge, & Buckolz, 2009; Schlaghecken & Sisman, 2006) and would be triggered differently (Schlaghecken et al., 2007) for masked and nonmasked distractors, self-inhibition may generate consistent inhibitory aftereffects whether its participation is invoked by visible or invisible primes or in identity or location tasks.
We wonder whether our masked target data represents a small departure from the extremely interesting view that subliminal processing is flexible, showing an ability to adapt to changes in task procedures that are put into practice on probe trials in response to visible stimuli (NCE tasks; e.g., Eimer & Schlaghecken, 1998; Klapp & Haas, 2005; O’Connor & Neill, in press). For example, O’Connor and Neill used compatible and incompatible stimulus–response mappings in a classic NCE task. They reported that the response manifesting an NCE was the one assigned to the presented event by the experimenter, showing that subliminal processing utilized whatever stimulus–response association rules were in effect. While this has yet to be shown for SNP tasks, we note that introducing free choice trials into the SNP procedure had the same outcome for both masked and nonmasked distractor primes. In any event, the subliminal target processing used the location–response assignments practiced on the probe trial (i.e., the target’s related response manifested the inhibitory aftereffects). However, the location–response rule practiced on the probe trial was not fully applied, because the target response was inhibited rather than executed. This, of course, is a result of the mask not just preserving prime event invisibility, but participating in probe aftereffect production. Hence, the subliminal processing of target events would not fully reflect their probe-trial processing.
Free choice response selection determinants
The study of free choice behavior warrants attention, because it occurs relatively frequently in everyday experience yet is underrepresented in the literature. With respect to isolating the factors that control “free choice,” we can now include recently executed responses, which are selected for, and ER, which causes an avoidance of recently inhibited responses. We also verified that hand preference is a strong selection determinant and will obscure ER manifestation (Appendix A), a problem that had been predicted due to the Buckolz et al. (2004) study. Efforts to examine ER should bear this in mind.
Notes
The distractor for the present experiment served as the target object for the pilot study, which used only prime trials. Prime distractor offset to mask onset delays of 0, 30, and 100 ms occurred randomly. The participants were instructed to respond only when a prime event was detected. Response rates to the target presentations were 11.5%, 85.3%, and 95.7%, for the 0-, 30-, and 100-ms delays, respectively. The low response rate with the 0-ms delay suggested that the mask largely maintained the phenomenal invisibility of the briefly presented prime event (i.e., the main experiment’s distractor event). This high rate of invisibility was reinforced by participants’ verbal reports.
References
Allport, D. A., Tipper, S. P., & Chmiel, N. R. J. (1985). Perceptual integration and postcategorical filtering. In M. I. Posner & O. S. M. Marin (Eds.), Attention and performance XI (pp. 107–132). Hillsdale, NJ: Erlbaum.
Baylis, G. C., Tipper, S. P., & Houghton, G. (1997). Externally cued and internally generated selection: Differences in distractor analysis and inhibition. Journal of Experimental Psychology. Human Perception and Performance, 23, 1617–1630.
Berlucchi, G. (2006). Inhibition of return: A phenomenon in search of a mechanism and a better name. Cognitive Neuropsychology, 23, 1065–1074.
Brown, M., Almeida, Q., Fitzgeorge, L., Buckolz, E. (2009). Is the basal ganglia involved in unconscious priming? Direct comparison of conscious and unconscious processing in individuals with Parkinson’s disease. Paper presented at the XVIII WFN World Congress on Parkinson’s Disease and Related Disorders, Miami Beach, FL.
Buckolz, E., Boulougouris, A., & Khan, M. (2002). The influence of probe-trial selection requirements on the location negative priming effect. Canadian Journal of Experimental Psychology, 56, 274–283.
Buckolz, E., Goldfarb, A., & Khan, M. (2004). The use of a distractor-assigned response slows later responding in a location negative priming task. Perception & Psychophysics, 66, 837–845.
Buckolz, E., Avramidis, C., & Fitzgeorge, L. (2008). Prime-trial processing demands and their impact on distractor processing in a spatial negative priming task. Psychological Research, 72, 235–248.
Buckolz, E., Fitzgeorge, L., Knowles, S. (2011). Spatial negative priming, but not inhibition of return, with central (foveal) displays. Manuscript under review.
Chao, H. F. (2009). Revisiting the prime-probe contextual similarity effect on negative priming: The impact of cue variability. European Journal of Cognitive Psychology, 21, 484–500.
Chao, H. F. (2011). Dissociations between identity and location negative priming. Acta Psychologica, 136, 81–89. doi:10.1016/j.actpsy.2010.10.005
Christie, J., & Klein, R. (2008). On finding negative priming from distractors. Psychonomic Bulletin & Review, 15, 866–873.
De Jong, R., Liang, C.-C., & Lauber, E. (1994). Conditional and unconditional automaticity: A dual-process model of effects of spatial stimulus-response correspondence. Journal of Experimental Psychology. Human Perception and Performance, 20, 731–750.
Eimer, M., & Schlaghecken, F. (1998). Effects of masked stimuli on motor activation: Behavioral and electrophysiological evidence. Journal of Experimental Psychology. Human Perception and Performance, 24, 1737–1747.
Eimer, M., & Schlaghecken, F. (2002). Links between conscious awareness and response inhibition: Evidence from masked priming. Psychonomic Bulletin & Review, 9, 514–520.
Eriksen, C. W., Coles, M. G., Morris, L. R., & O’Hara, W. P. (1985). An electromyographic examination of response competition. Bulletin of the Psychonomic Society, 23, 165–168.
Fitzgeorge, L., & Buckolz, E. (2008). Spatial negative priming modulation: The influence of probe-trial target cueing, distractor presence and an intervening response. European Journal of Cognitive Psychology, 20, 994–1026.
Fitzgeorge, L., & Buckolz, E. (2009). Automatic versus volitional orienting and the production of the inhibition-of-return effect. Canadian Journal of Experimental Psychology, 63, 94–102.
Gibbons, H., & Rammsayer, T. H. (2004). Differential effects of prime–probe duration on positive and negative location priming: Evidence for opponent facilitatory and inhibitory influences in priming tasks. The Quarterly Journal of Experimental Psychology, 57A, 61–86.
Guy, S., & Buckolz, E. (2007). The locus and modulation of the location negative priming effect. Psychological Research, 71, 178–191.
Guy, S., Buckolz, E., & Pratt, J. (2004). The influence of distractor-only prime trials on the location negative priming mechanism. Experimental Psychology, 51, 4–14.
Guy, S., Buckolz, E., & Khan, M. (2006). The locus of location repetition latency effects. Canadian Journal of Experimental Psychology, 60, 307–318.
Guy, S., Buckolz, E., & Fitzgeorge, L. (2007). Disengaging the location negative priming effect: The influence of an intervening response. European Journal of Cognitive Psychology, 19, 789–812.
Harvey, N. (1980). Non-informative effects of stimuli functioning as cues. The Quarterly Journal of Experimental Psychology, 32, 413–425.
Hommel, B. (1993). The role of attention for the Simon effect. Psychological Research, 55, 208–222.
Klapp, S. T., & Haas, B. W. (2005). Nonconscious influence of masked stimuli on response selection is limited to concrete stimulus–response associations. Journal of Experimental Psychology. Human Perception and Performance, 31, 193–209.
Klapp, S. T., & Hinkley, L. B. (2002). The negative compatibility effect: Unconscious inhibition influences reaction time and response selection. Journal of Experimental Psychology. General, 131, 255–269.
Klein, R. M. (2000). Inhibition of return. Trends in Cognitive Sciences, 4, 138–147.
Klein, R., Christie, J., & Morris, E. P. (2005). Vector averaging of inhibition of return. Psychonomic Bulletin & Review, 12, 295–300.
Mayr, S., Hauke, R., & Buchner, A. (2009). Auditory location negative priming: A case of feature mismatch. Psychonomic Bulletin & Review, 16, 845–849.
McCormick, P. A. (1997). Orienting attention without awareness. Journal of Experimental Psychology. Human Perception and Performance, 23, 168–180.
Milliken, B., Tipper, S. P., Houghton, G., & Lupiáñez, J. (2000). Attending, ignoring, and repetition: On the relation between negative priming and inhibition of return. Perception & Psychophysics, 62, 1289–1296.
Moore, C. (1994). Negative priming depends on probe-trial conflict. Perception & Psychophysics, 56, 133–147.
Mulckhuyse, M., & Theeuwes, J. (2010). Unconscious attentional orienting to exogenous cues: A review of the literature. Acta Psychologica, 134, 299–309.
Neill, W. T. (2007). Mechanisms of transfer-inappropriate processing. In D. S. Gorfein & C. M. MacLeod (Eds.), Inhibition in cognition (pp. 63–78). Washington, DC: American Psychological Association.
Neill, W. T., Terry, K. M., & Valdes, L. A. (1994). Negative priming without probe selection. Psychonomic Bulletin & Review, 1, 119–121.
Neill, W. T., Valdes, L. A., & Terry, K. M. (1995). Selective attention and the inhibitory control of cognition. In F. N. Dempster & C. J. Brainerd (Eds.), Interference and inhibition in cognition (pp. 207–255). San Diego: Academic Press.
O’Connor, P. A., & Neill, W. T. (in press). Does subliminal priming of free response choices depend on task set or automatic response activation? Consciousness and Cognition. doi:10.1016/j.concog.2010.08.007
Park, J., & Kanwisher, N. (1994). Negative priming for spatial locations: Identity mismatching, not distractor inhibition. Journal of Experimental Psychology. Human Perception and Performance, 20, 613–623.
Rafal, R., Calabresi, P., Brennan, C., & Sciolto, T. (1989). Saccade preparation inhibits reorienting to recently attended locations. Journal of Experimental Psychology. Human Perception and Performance, 15, 673–685.
Rafal, R., Davies, J., & Lauder, J. (2006). Inhibitory tagging at subsequently fixated locations: Generation of “inhibition of return” without saccade inhibition. Visual Cognition, 13, 308–323.
Schlaghecken, F., & Eimer, M. (2004). Masked prime stimuli can bias “free” choice between response alternatives. Psychonomic Bulletin & Review, 11, 463–468.
Schlaghecken, F., & Sisman, R. (2006). Subliminal response priming in children: Evidence for a subcortical locus of the negative compatibility effect. Advances in Cognitive Psychology, 2, 7–19.
Schlaghecken, F., Rowley, L., Sembi, S., Simmons, R., & Whitcomb, D. (2007). The negative compatibility effect: A case for self-inhibition. Advances in Cognitive Psychology, 3, 227–240.
Schlaghecken, F., Klapp, S. T., & Maylor, E. A. (2009). Either or neither, but not both: Locating the effects of masked primes. Proceedings of the Royal Society B, 276, 515–521.
Sumner, P. (2007). Negative and positive masked-priming—implications for motor inhibition. Advances in Cognitive Psychology, 3, 317–326.
Valle-Inclan, F., & Redondo, M. (1998). On the automaticity of ipsilateral response activation in the Simon effect. Psychophysiology, 35, 366–371.
Van der Stigchel. (2010). Recent advances in the study of saccade trajectory deviations. Vision Research, 50, 1619–1627.
Vaughan, F. L., Hughes, E. A., Jones, R. S. P., Woods, R. T., & Tipper, S. P. (2006). Spatial negative priming in early Alzheimer’s disease: Evidence for reduced cognitive inhibition. Journal of the International Neuropsychological Society, 12, 416–423.
Welsh, T. N. (2002). The influence of response priming, competition, and inhibition on movement trajectories. Unpublished PhD thesis, McMaster University, Hamilton, Ontario, Canada.
Acknowledgements
This work was supported by an operating grant to E.B. and a doctoral scholarship to L.F. from the Natural Sciences and Engineering Research Council of Canada. Thanks to Ioan Todoran for programming this experiment.
Author information
Authors and Affiliations
Corresponding author
Appendices
Appendix A
Appendix B. Different-hand competitions (free choice trials)
Distractor primes
Results
Different-hand competitions: CO versus CO
A significant chi-squared value emerged, χ 2(1, n = 1088) = 44.49, p < .05, for different-hand finger competitions involving control responses. Follow-up tests showed that the preferred-hand finger alternative was selected significantly more often than the nonpreferred option for both the no-mask, χ 2(1, n= 272) = 158.89, p < .05, and masked, χ 2(1, n = 272)= 14.24, p < .05, primes. This imbalance was reliably larger with no-mask than with masked primes (.77/.23 vs. .58/.42).
Different-hand competitions: DRR versus CO
With competing DRR versus CO response types, both inhibition avoidance and hand preference were free to operate as response selection determinants. For the nonmasked primes, the chi-squared value was nonsignificant, χ 2(1, n = 544) = 0.069, p > .05. Follow-up tests revealed that hand preference, χ 2(1, n=544)=113.06, p < .05, but not response type, χ 2(1, n = 544) = 0.265, p > .05, influenced free choice response selection.
In contrast, with masked prime distractor locations, a significant chi-squared value, χ 2(1, n=544)=4.15, p < .05, pointed to a combined impact of hand preference and response type on response selection. Both factors exerted a significant influence on response choice. The preferred-hand finger response, χ 2(1, n=544)=26.47, p < .05, and the control type of response, χ 2(1, n=544)=70.62, p < .05, were used more often than their respective alternatives overall. The interactive impact of these two selection determinants was evident in various ways. For example, the prime distractor response was more likely chosen if it was associated with the preferred-hand (.43) than with the nonpreferred-hand (.21) option. Alternatively, when the preferred-hand finger response had previously been inhibited, it was chosen less often (.43) than the nonpreferred-hand finger response that had not undergone inhibition (.57).
Reaction time
Reliable aftereffect and mask type main effects were qualified by their significant interaction, F(1, 16)=8.95, MSE=1,254, p < .01. Post-hoc tests (p < .01) showed that the inhibitory aftereffect was present for both nonmasked (69 ms) and masked (32 ms) primes, but was significantly larger in the former case.
Discussion
When the allowable finger responses on free-choice trials were on different hands following a nonmasked prime distractor event, the preferred-hand finger response was chosen significantly more often (about 75%) than the alternative, and this selection preference was indifferent to whether the finger had recently been inhibited or not (DRR vs. CO). Thus, as expected (Buckolz et al., 2004), different-hand competitions provided no evidence for an execution resistance property attendant on recently inhibited (DRR) responses. In light of this result and of our same-hand competition data, different-hand free choice competitions should not be used to study the manifestation of execution resistance on response selection. Finally, while execution resistance was not of sufficient relative strength to influence response selection in free choice, it nonetheless still produced its characteristic RT delay when an inhibited response was actually chosen (inhibitory aftereffect).
The impact of masking the prime distractor event produced aftereffect changes for different-hand competitions that were similar to those seen for same-hand competitions; namely, hand preference exerted significantly reduced control over free choice response selection, and the size of the inhibitory aftereffect decreased significantly with a mask (69 vs. 32 ms). These results may reflect the operation of the self-inhibition operation suggested by Schlaghecken et al. (2007) as being triggered by a mask.
Target primes
Results
Different-hand competitions: CO versus CO
When the competing fingers were both of the control variety, a significant chi-squared value emerged, χ 2(1, n = 1088) = 16.70, p < .05. The preferred-hand finger response was used significantly more often than the control alternative for both the no-mask, χ 2(1, n = 544) = 56.94, p < .05, and masked, χ 2(1, n = 544) = 3.56, p < .05, primes, with this imbalance being reliably larger in the former instance (.66/.34 vs. .54/.46). This pattern is reminiscent of the one seen with prime distractor locations (see Table 1).
Different-hand competitions: TRR versus CO
Chi-squared values were calculated for no-mask and masked primes separately, with Hand Preference and Response Type as the main factors. When no mask was used, the chi-squared value was nonsignificant, χ 2(1, n=544)= 1.83, p > .05. Follow-up tests revealed that both factors significantly influenced response choice; preferred-hand finger responses χ 2(1, n = 544) = 90.60, p < .05, and primed target outputs, χ 2(1, n= 544) = 9.07, p < .05, were selected more often than their competitors. When prime target events were masked, the chi-squared value approached significance, χ 2(1, n = 544) = 3.53, p = .06. Follow-up tests indicated that, overall, the preferred-hand finger response (.72/.28) and the control response type (.58/.42) were chosen significantly more often than their respective alternatives.
Reaction time
A significant aftereffect main effect, F(1, 16) = 23.07, MSE = 1,632, p < .01, was qualified by its reliable interaction with mask type, F(1, 16) = 70.56, MSE = 2,203, p < .01. The aftereffect was positive following a nonmasked prime (−101 ms) but negative following a masked prime (+35 ms).
Discussion
When the target primes were not masked, and when aftereffect influences could not affect response choice (CO vs. CO), the preferred-hand finger was selected significantly more often than the nonpreferred-hand finger. This same bias occurred when both aftereffect and hand preference were free to operate as response selection determinants (TRR vs. CO), and while this inclination was greater when the preferred hand response had been executed on the prime trial (.77 vs. .64), it was not a significant difference. As with the nonmasked distractor primes, possible aftereffects did not influence response selection for different-hand finger response competitions.
The preferred-hand bias was significantly smaller with masked than with nonmasked target primes, for both CO versus CO and TRR versus CO free choice competitions. In the latter instance, the preferred-hand response was chosen less often (approaching significance) than when it served as the target response on the prime (.64 vs. .80), presumably because it had just undergone inhibition. Hence, we see an impact of execution resistance on free choice response selection for masked target primes. This is supported by the fact that reactions for masked target-repeat trials were now significantly slower than control reactions (31 ms), demonstrating an inhibitory aftereffect, as predicted by the self-inhibition mechanism (Schlaghecken et al., 2007).
Rights and permissions
About this article
Cite this article
Fitzgeorge, L., Buckolz, E. & Khan, M. Recently inhibited responses are avoided for both masked and nonmasked primes in a spatial negative priming task. Atten Percept Psychophys 73, 1435–1452 (2011). https://doi.org/10.3758/s13414-011-0125-7
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13414-011-0125-7