
Abstract
Response-irrelevant stimuli can be encoded with, and later on retrieve, a response given to a relevant stimulus, an effect that is called distractor–response binding. In three experiments using a prime–probe design, we investigated whether the allocation of attention modulates the processes contributing to distractor–response binding. Participants identified letters via keypresses while attending to one of two sets of simultaneously presented but response-irrelevant number stimuli. In different experiments, both spatial attention and feature-based attention were allocated to the response-irrelevant stimuli. The results showed that only attended response-irrelevant stimuli elicited effects of distractor–response binding. In particular, while the encoding of response-irrelevant stimuli and responses was not particularly affected by attention during prime processing, only attended response-irrelevant stimuli in the probe retrieved previous responses. Hence, we show that attention affects action regulation due to modulating the influence of stimulus–response binding on behavior.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Two mechanisms that (among others) allow for efficient orienting and responding in complex environments are attention and the integration of stimuli with executed responses. Selective attention allows for attending and responding to only those aspects of our environment that are related to our goals and ensures that we are not distracted by stimuli that are irrelevant for or might interfere with the current task. Thus, selective attention is important for the top-down control of behavior (Tipper, 1992). However, not all our actions are intentionally controlled. Obviously, we have other mechanisms that translate (originally) intentional actions into efficient behavioral routines and habits. The retrieval of previous integrated stimuli and responses might play an important role for such an automatization of behavior (e.g., Logan, 1988). Yet, it is not clear, what role attentional mechanisms play in the early stages of association development. In this article, we therefore focus on the influence selective attention might have on the integration and retrieval of single stimulus–response episodes.
For stimuli that are encountered more than once, it is adaptive to form a direct association between the stimulus and the (successful) response. It has been proposed that each encounter of a stimulus is encoded in an instance (Logan, 1988, 1990) or event file (Hommel, 2004; Hommel, Müsseler, Aschersleben, & Prinz, 2001) that also includes the action performed in response to the stimulus. With the accumulation of instances of a certain situation, fast memory-based retrieval of the earlier actions can replace the algorithmic processing of the response indicated by the stimulus. A considerable amount of evidence exists that indicates temporary stimulus–response binding after a single encounter as well as the formation of long-term associations between stimuli and responses due to the repeated encounter of similar events (e.g., Boronat & Logan, 1997; Hommel & Colzato, 2004, 2009; Lassaline & Logan, 1993; Logan, 1992; Logan, Taylor, & Etherton, 1996, 1999). More specifically, it has been shown that not only stimuli that people respond to but also response-irrelevant stimuli can become temporarily integrated with responses in a single encounter and thus trigger response retrieval later on, a phenomenon labeled distractor–response binding (Rothermund, Wentura, & De Houwer, 2005; see also Frings, Rothermund, & Wentura, 2007). Whether binding of stimuli and responses can become an action routine that can be retrieved from memory is controlled through several feedback mechanisms. For example, event files that trigger erroneous responses are easily detected by our cognitive system (Wiswede, Rothermund, & Frings, 2013). In addition, event files that received negative feedback have been shown to be deleted from memory (e.g., Rothermund, Eder, & Frings, 2014; Waszak & Pholulamdeth, 2009). In sum, binding between (ir) relevant stimuli and responses can be understood as a mechanism in the early stages of routine development in human action control.
The influence of attention on the integration of stimulus features and on the integration of stimulus and response features
Responding to certain stimuli is typically preceded by orienting to and identification of the relevant stimulus. For example, to pick up an apple from a bowl of fruit, a person would have to find out whether there is an apple left in the bowl, attend to the apple while ignoring other fruit lying next to it, and finally start the grasping movement. That is, before an action is possible, a person oftentimes has to identify whether an object carrying certain features is present in a given situation. In order to identify an object that is defined by more than one feature, the feature integration theory of attention proposes that (spatial) attention has to be focused on the stimulus’s location, leading to a binding of all stimulus features into an object file (Kahneman, Treisman, & Gibbs, 1992; Treisman 1996; Treisman & Gelade, 1980). Thus, spatial attention seems to be a precondition for feature binding (see also Engel & Singer, 2001; Treisman, 1996; von der Malsburg, 1999). In addition, attention might influence not only the binding of stimulus features into objects, but also the integration of stimulus and response features (see Hommel et al., 2001; Logan, 1988, 1990; Memelink & Hommel, 2012). For example, Logan (1988, 1990) proposes that attention obligatorily and unavoidably leads to encoding of the attended stimulus into memory, and that the same act of attention obligatorily and unavoidably retrieves from memory what has been associated with that stimulus—that is, also associated responses. Put differently, it can be assumed that people learn those co-occurrences they attend to (Logan & Etherton, 1994). Similarly, the theory of event coding (Hommel et al., 2001), suggests that intentional weighting (i.e., a process very similar to the concept of attention, see Logan, 2002; Müller, Heller, & Ziegler, 1995) plays an important role for event coding (Memelink & Hommel, 2012). In fact, attention to response-irrelevant stimuli increased the strength of stimulus–response associations that were learned during repeated encounters of constant stimulus–response combinations in several studies (Boronat & Logan, 1997; Logan, 1998; Logan & Etherton, 1994; Logan et al., 1996, 1999).
In addition, it has been shown that attentional processes also facilitate binding between stimulus features after a single encounter of a stimulus–response combination (Hommel & Colzato, 2009; van Dam & Hommel, 2010). However, no clear evidence exists regarding the binding of stimulus features and responses in the early stages of association formation (i.e., after a single encounter). In fact, several studies suggested that the likelihood of attention being deployed to stimuli does not modulate stimulus–response binding processes at all (e.g., Hommel, 2005, 2007; Hommel & Colzato, 2004). Yet, the integrated features in the studies investigating stimulus–response bindings were typically part of the target stimuli and varied on a task relevant dimension. These conditions might have ensured that some attention was allocated to the stimulus features in all cases, which may have been sufficient for features to be integrated with responses. Thus, so far it cannot be ruled out that attention is also a prerequisite for temporary stimulus–response bindings.
Binding effects are typically measured in sequences of two responses that are triggered by targets in a stimulus arrangement. Stimulus-features are integrated with the first response and can retrieve it, if they are repeated in the second stimulus arrangement. If the second response is the same as the first, such retrieval enhances performance. In contrast, if the first and second responses differ, response retrieval due to stimulus repetition hinders performance. Hence, binding is indicated by facilitation and interference effects in response times and error rates of the second response in such sequences. Participants by definition attend to target objects. And processing of one feature of an object can be assumed to automatically open an attentional gate to the processing of other features of this object (e.g., Baylis & Driver, 1992, 1993; Duncan, 1984). Therefore, to investigate whether attention plays a crucial role in bindings of stimuli and responses, additional task-irrelevant stimuli are necessary. As we mentioned above, the effect of distractor–response binding indicates that additional stimuli that are irrelevant for the current task can also be integrated with the response and retrieve it later on (see, e.g., Frings, 2011; Frings & Moeller, 2010; Frings, Moeller, & Rothermund, 2013; Frings & Rothermund, 2011; Giesen, Frings, & Rothermund, 2012; Giesen & Rothermund, 2011, 2013; Moeller & Frings, 2011; Moeller, Rothermund, & Frings, 2012). Investigating the role attention plays for this mechanism would be a purer test of whether attention is vital for stimulus–response bindings to influence behavior.
Giesen et al. (2012) used targets (letters) and distractors (one digit numbers) that did not belong to the same stimulus set. Hence, distractors likely received less attention than distractors in studies that used the same four letters both as targets and distractors (e.g., Frings & Moeller, 2012). Nevertheless, presenting targets and distractors in a horizontal arrangement, Giesen and colleagues found evidence that distractor–response binding is generally possible when distractors are drawn from a different set of stimuli than targets. However, additional stimuli were not always automatically integrated in previous studies. In particular, the arrangement of target and distractor stimuli influenced whether or not distractors were integrated with and later on retrieved responses to targets (Frings & Rothermund, 2011; Giesen & Rothermund, 2011; Moeller et al., 2012). Frings and Rothermund concluded that grouping principles modulate binding processes: If target and distractor are presented in a grouped arrangement, binding of distractors and responses takes place. In contrast, if target and distractor are presented in a non-grouped setup the effect of distractor–response binding is significantly smaller or does not occur at all. Yet, in Experiments 1–5 of their study, grouping manipulations were confounded with differences in spatial distance. In turn, spatial attention might have influenced distractor–response binding in these experiments, as well. With the present series of experiments, we aimed to systematically test the influence attentional distribution has on the integration of responses with stimuli that are entirely irrelevant for the required response.
Overview
On an abstract level, we examined whether instructed focus of attention has an effect on binding processes regarding stimuli that are entirely response-irrelevant. If participants were instructed to attend to some of the response-irrelevant stimuli, we assumed that distractor–response binding effects would occur only for the attended, but not for the unattended response-irrelevant stimuli. In a first step we manipulated the likelihood of attention being distributed to the distractor stimuli, to investigate whether attention can modulate binding of response-irrelevant stimuli and responses. Then, we tested whether different kinds of attention (namely, spatial and feature-based attention) influence binding processes differently. Finally, we analyzed whether the integration of response and stimulus features or the distractor-based retrieval of responses was affected by the allocation of attention.
More concretely, we analyzed the effect of distractor–response binding in a paradigm including two subsequent forced choice responses (i.e., a prime–probe design) in which response relation (repetition vs. change) and distractor identity relation (repetition vs. change) are varied orthogonally. In this paradigm, evidence for distractor–response binding can be seen if performance is better due to repeated distractors if responses repeat, but impaired due to repeated distractors if responses change (i.e., the Distractor Relation × Response Relation interaction taps the effect of distractor–response binding). In all experiments reported here, participants’ task was to identify a target letter (D, F, J, or K) that was presented in the center of the display by pressing the corresponding key. Two sets of additional distractor stimuli were simultaneously presented with each target letter. Each distractor set consisted of two identical digits that were arranged in diagonal locations adjacent to the target letter (i.e., upper left/ lower right and upper right/ lower left positions). To vary the likelihood of attention being distributed to different distractor stimuli, participants were instructed to attend to one set of distractor stimuli, whereas the other distractor set could be ignored. After a third of all prime–probe sequences, participants were asked to report the identities of the to-be-attended distractors that had been presented on the prime and on the probe display. None of the distractor stimuli, regardless of whether they were to be attended or unattended, were relevant for the responses to the target stimuli. In Experiment 1, attention was drawn to two of the four distractor positions on a spatial basis, via instruction or visual cues. Assuming spatial control of attention (e.g., Posner, Snyder, & Davidson, 1980), the spatial cues would enable participants to allocate their attention to the relevant locations before stimulus onset. In Experiment 2, participants selected to-be-attended distractors via features independent of the distractors’ locations. Thus, participants had to visually search for the relevant distractor, which ensured that to-be-unattended distractors (sharing features with the relevant distractors) would also receive some attention. Finally, in Experiment 3, we varied whether prime or probe distractors were to be attended in order to analyze whether integration or retrieval processes were modulated by attention. On the basis of earlier studies on distractor–response binding effects, we expected to find large- to medium-sized effects, and planned the sample sizes of each experiment accordingly.
Experiment 1a
Experiment 1a investigated whether it is possible to modulate the effect of distractor–response binding by the attention focused on the distractor stimuli. Since spatial attention can modulate integration of stimulus features (van Dam & Hommel, 2010), our first attempt to modulate distractor–response binding was to vary the probability of attention being focused on locations of these distractors.
We used a stimulus arrangement, including four distractor stimuli and one target, and varied the likelihood of attention being directed to the response-irrelevant stimuli. Participants’ task was to identify a centrally presented target letter by pressing an according key. Four task-irrelevant distractor stimuli (one digit numbers) appeared in positions surrounding the target letter. Targets and distractors were drawn from different stimulus sets to minimize the response relevance of the additional stimuli. Participants were instructed to always attend to two of the distractor positions and to completely ignore the other distractor locations. Hence, distractor–response binding effects could be measured separately for distractors appearing in the to-be-attended and to-be-unattended distractor positions. The to-be-attended and to-be-unattended distractors were always drawn from different stimulus sets (the digits 1–4 and 5–8, respectively). Consequently, the to-be-unattended distractors not only always appeared at irrelevant locations, but their identities were also never to be attended during the experiment. If attention has no influence on the integration and retrieval of event files (see Hommel, 2005, 2007), we expected to find the same result pattern, regarding distractor–response binding effects, for to-be-attended and to-be-unattended distractors. However, if attention to a stimulus influences its binding to a response, we would find different effects of distractor–response binding for to-be-attended and to-be-unattended distractors. Specifically, we expected no distractor–response binding effect for to-be-unattended distractors, but a significant effect of distractor–response binding for to-be-attended distractors, in Experiment 1a. In the present design, in which we orthogonally varied response relation and distractor relation, distractor–response binding effects would be statistically evidenced by an interaction of response relation and distractor relation: If the response had to be repeated, response retrieval due to a repeated distractor should enhance performance. That is, for response repetition trials, faster responses were predicted for distractor repetition than for distractor change trials. In contrast, in response change trials, a repeated distractor would trigger the retrieval of an inadequate response. Therefore, for response change trials less advantage due to distractor repetition would be expected than in response repetition trials. In some cases, distractor repetition in response change trials would even lead to worse performance than distractor change. Thus, if attention modulates distractor–response binding, we expected to see an interaction of response relation and distractor relation for to-be-attended distractors and a significantly smaller, or no, interaction if distractors were to be unattended.
Method
Participants
A group of 24 students (13 women, 11 men) from the University of Trier took part in the experiment. Their median age was 22 years with a range from 19 to 28 years. All participants took part in exchange for partial course credit.
Design
The design of Experiment 1a comprised three within-subjects factors, namely response relation (repetition vs. change), distractor relation (repetition vs. change), and instructed attention to the distractors (distractors to be attended vs. unattended).
Materials
The experiment was conducted using the E-Prime software (version 2.0). Instructions and all stimuli were shown in white on black background on a standard CRT screen. The stimuli were the letters D, F, J, and K and the digits 1–8. In one third of the trials, participants were prompted to report the identity of the digits on the prime and the probe in one distractor set via two red question marks that were presented after the probe response. All letters and digits as well as the question marks were 0.8 to 1.1 cm wide and 0.9 to 1.0 cm high. The entire arrangement of five characters had a horizontal and vertical distribution of 2.8 cm.
Procedure
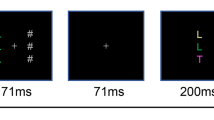
Participants were tested individually in soundproof chambers. Instructions were given on the screen and summarized by the experimenter. Participants were instructed to place the index and middle fingers of their hands on the keys D, F, J, and K of a standard computer keyboard. Their task was always to identify the target letter by pressing a key with the corresponding finger. Each prime and each probe display included one target letter (D, F, J, or K) and four distractor digits that were arranged around the target at the corners of an imagined square. The digits on each diagonal had the same identity, forming one set of distractors on each diagonal. To minimize attention that might be drawn to the to-be-unattended distractor set, the to-be-attended and to-be-unattended distractors were always drawn from different digit pools and always presented in constant “to-be-attended” and “to-be-unattended” locations. Hence, on each prime and each probe, one of the distractor identities was 1, 2, 3, or 4, and the other identity was 5, 6, 7, or 8. Half of the participants saw the digits 1–4 in the upper right and lower left positions and the digits 5–8 in the upper left and lower right positions, whereas this mapping was reversed for the other half of the participants. Orthogonally to this mapping, half of the participants were instructed to attend to the distractor digits in the upper left and lower right positions and the other half was instructed to attend to the digits in the upper right and lower left positions. Participants were instructed to react as quickly and as correctly as possible. A single trial (prime–probe sequence) consisted of the following sequence of events (see Fig. 1a): Participants started each trial by pressing the space bar. After 500 ms of a blank screen, the prime display appeared and stayed on the screen until participants responded. After the prime response, another blank display of 500-ms duration appeared, before the probe display was presented. As with the prime, the probe target and distractors stayed on the screen until participants responded. On two thirds of the trials, the probe response was followed by 500 ms of a blank screen before an asterisk appeared, indicating that the next trial could be started. To ensure that participants attended to one set of distractors and ignored the other, on one third of the trials the probe response was followed by a prompt, asking which identities the to-be-attended distractors had on the second to last (i.e., the prime) and the last (i.e., the probe) displays. In response repetition trials, the same letter was presented as the target on the prime and on the probe, whereas on response change trials, the target identity changed from prime to probe. Orthogonally to the response relation, the distractor relation was varied. In distractor repetition trials, the prime distractor identity was repeated as the probe distractor identity, whereas in distractor change trials, the distractor identity changed from prime to probe. On half of the trials, distractor repetition or change referred to the to-be-attended distractor set, and on the other half, it referred to the to-be-unattended distractor set. To ensure that any effects of retrieval were entirely due to either the to-be-attended or to-be-unattended distractors, the distractor in the other set always changed from prime to probe. For example, if a trial included distractor repetition of the to-be-unattended distractor, the identity of the to-be-attended distractor changed from prime to probe. In turn, four different conditions were created, for both to-be-attended and to-be-unattended distractors: In response repetition/distractor repetition trials, the prime target and the prime distractor were repeated on the probe. In response repetition/distractor change trials, the prime target was repeated as the probe target and the distractor changed from prime to probe. In response change/distractor repetition trials, the probe target differed from the prime target and the prime distractor was repeated as the probe distractor. Finally, in response change/distractor change trials, no stimulus was repeated from the prime to the probe display.

Upper panel: Sequence of events in one trial in Experiments 1a and 3. For participants instructed to attend to the upper left and lower right distractors, this is an example of a response repetition, to-be-attended distractor repetition trial; for participants instructed to attend to the upper right and lower left distractors, it is an example of a response repetition, to-be-unattended distractor repetition trial. Lower panel: Sequence of events in one trial in Experiment 1b. Participants were informed, via the locations of two exclamation marks before each prime and each probe, which distractor locations had to be attended in the following display. The example illustrates a response repetition/distractor repetition trial for the to-be-attended distractors. On a third of the trials, two displays were presented after the probe response, prompting participants to report the identities of the to-be-attended distractors. On two thirds of the trials, these two displays were replaced by 500 ms of a blank screen. Stimuli are not drawn to scale
Each participant worked through one experimental block of 192 prime–probe sequences. The four trial types (response repetition/distractor repetition, response repetition/distractor change, response change/distractor repetition, and response change/distractor change) were realized in 24 trials each for the to-be-attended and to-be-unattended distractors. Two of the letters D, F, J, and K were randomly assigned to be the prime target and probe target. Similarly, the to-be-attended prime and probe distractors were drawn from one digit pool and the to-be-unattended distractors were drawn from another pool. For each trial sequence, stimuli were then changed in accordance with the particular condition; for example, in an attended response change/distractor repetition trial, the identity of the to-be-attended prime distractor was then changed to the identity of the to-be-attended probe distractor. Before the experimental block started, participants worked through a practice block of 32 prime–probe sequences in which they received feedback after each response. Participants were randomly prompted after half of these sequences to report the identities of the to-be-attended prime and probe distractors. At the beginning of the practice block, four additional prime–probe sequences were presented, after which participants were always asked to report the identities of the to-be-attended prime and probe distractors. Everything else was exactly as during the experimental trials.
Results
Only trials with correct answers to both the prime and the probe were considered. Reaction times that were more than 1.5 interquartile ranges above the third quartile of a participant’s distribution (Tukey, 1977), and those that were shorter than 200 ms, were excluded from the analysis. Due to these constraints, 8.3 % of all trials were discarded (the probe error rate was 2.2 %, and the prime error rate was 2.7 %). The mean reaction times and error rates for probe displays are depicted in Table 1. The significance criterion for all reported analyses was set to p < .05 (two tailed).
Reaction times
In a 2 (response relation: response repeated vs. response changed) × 2 (distractor relation: distractor repeated vs. distractor changed) × 2 (instructed attention: distractors to be attended vs. unattended) multivariate analysis of variance (MANOVA) with Pillai’s trace as the criterion, all main effects of response relation, distractor relation, and instructed attention were significant: F(1, 23) = 79.33, p < .001, ηp 2 = .78, for response relation; F(1, 23) = 12.00, p = .002, ηp 2 = .34, for distractor relation; and F(1, 23) = 9.06, p = .006, ηp 2 = .28, for instructed attention. Responses to repeated targets (M = 600 ms, SD = 117 ms) were faster than those to changed targets (M = 843 ms, SD = 194 ms); responses were faster if distractors were repeated (M = 704 ms, SD = 136 ms) than if they changed from prime to probe (M = 728 ms, SD = 152 ms); and responses were faster if the potentially repeated distractors were to be attended (M = 704 ms, SD = 133 ms) rather than to be unattended (M = 728 ms, SD = 155 ms). In addition, the interactions of Instructed Attention × Response Relation, F(1, 23) = 18.84, p < .001, ηp 2 = .45, and Instructed Attention × Distractor Relation, F(1, 23) = 9.08, p = .006, ηp 2 = .28, were both significant. The advantage of response repetition was larger if distractors were to be attended rather than unattended, and the advantage of repeated distractors was also larger if the distractors were to be attended rather than unattended. More interestingly, the interaction of response relation and distractor relation was significant, as well, F(1, 23) = 10.21, p = .004, ηp 2 = .31, indicating that distractor repetition led to a larger advantage in response repetition than in response change trials. Most importantly, the three-way interaction of instructed attention, response relation, and distractor relation was also significant, F(1, 23) = 14.94, p = .001, ηp 2 = .39. That is, the interaction of response relation and distractor relation was modulated by instructed attention to the distractors. Separate analyses revealed a significant interaction of response relation and distractor relation for to-be-attended distractors, F(1, 23) = 36.09, p < .001, ηp 2 = .61, but no interaction of response relation and distractor relation for to-be-unattended distractors, F(1, 23) = 1.71, p = .204, ηp 2 = .07.
Error rates
The same MANOVA on error rates revealed significant main effects of response relation, F(1, 23) = 36.36, p < .001, ηp 2 = .61, and instructed attention, F(1, 23) = 6.00, p = .022, ηp 2 = .21. Participants made more errors if responses changed from prime to probe than if the responses had to be repeated, and they made more errors if distractor repetition or change took place among the to-be-attended distractors rather than the to-be-unattended distractors. The interaction of response relation and instructed attention, F(1, 23) = 4.17, p = .053, ηp 2 = .15, as well as the three-way interaction of response relation, distractor relation, and instructed attention, F(1, 23) = 4.17, p = .053, ηp 2 = .15, did not reach significance. None of the other effects was significant, either, all Fs < 2.09, ps > .16. In separate analyses, the interaction of response relation and distractor relation was significant for neither to-be-attended distractors, F(1, 23) = 4.02, p = .058, ηp 2 = .15, nor to-be-unattended distractors, F(1, 23) = 0.52, p = .479, ηp 2 = .02.
Note that for categorical outcome variables, ANOVA can lead to spurious results; it has thus been suggested that researchers use mixed logit models instead (Jaeger, 2008). Thus, for this experiment and all further experiments, we will report the ANOVA for the error rates in the main text, but report multilevel logistic regression models in the Appendix.
Discussion
In Experiment 1a, the significant interaction of response relation and distractor relation indicated a general effect of distractor–response binding. Moreover, the interaction of response relation, distractor relation, and instructed attention indicated that the effect of distractor–response binding was modulated by instructed attention to the distractors. That is, to-be-attended distractors were integrated with and retrieved responses to a target stimulus, whereas to-be-unattended distractors did not.Footnote 1 This indicates that attention distributed to a distractor stimulus modulates whether or not this stimulus influences the processes of action control. Distractor–response binding only seems to occur if attention is focused on the distractor stimulus, but not if the distractor is unattended.
In line with studies regarding the automatization of behavior (e.g., Logan et al., 1999), the present results can be interpreted as confirming that unattended stimuli do not become part of instances: Even immediately following the formation of an instance, we found no indication that to-be-unattended stimuli had become part of the newly created instance.
Since the positions of to-be-attended and to-be-unattended distractors in Experiment 1a were always constant, it is possible that the to-be-attended distractor identity was not the only important element for retrieving prime responses, but that the repeated locations of the to-be-attended stimuli played an important role, as well. According to the feature integration theory of attention, objects are initially addressed via their location code (Kahneman & Treisman, 1984; Treisman, 1992, 1996). Yet, it has been shown that changing an object’s location from a study to a test display impeded object identification at short- (up to 1,000 ms) but not at longer (1,500–2,500 ms) study–test intervals, indicating a special role of stimulus location only for a short time span after encoding (Logie, Brockmole, & Jaswal, 2011). Also, Logie and colleagues investigated objects that were relevant for their participants’ task (i.e., target objects), whereas we were interested in the effect that the locations of response-irrelevant (i.e., distractor) objects have on encoding and retrieval processes. It can be assumed that the binding mechanisms of response-irrelevant stimuli have shorter durations than do bindings concerning target features (see Frings, 2011; Hommel & Colzato, 2004). Nevertheless, it is possible that the intervals between prime and probe were short enough in Experiment 1a that the repetition of distractor locations was crucial for the occurrence of distractor–response bindings. Experiment 1b was conducted to exclude this possibility.
Experiment 1b
The task and stimulus arrangements in Experiment 1b were identical to those used in Experiment 1a. However, in contrast to Experiment 1a, participants were not instructed to always attend to the same distractor positions and to completely ignore the other distractor locations. To be able to infer the effect of distractor–response binding for the to-be-attended distractors on distractor identity, independent of distractor location, in Experiment 1b the positions of the to-be-attended distractors varied randomly between trials and participants were informed about the relevant positions of the distractors before each display. Thus, all possible distractor positions included to-be-attended stimuli on half of the trials and to-be-unattended ones on the other half. We also allowed changes of to-be-attended and to-be-unattended distractor positions between the prime and probe responses. On half of the trials, the to-be-attended distractors were presented in the same locations on the prime and probe, and on the other half, the locations of the to-be-attended distractors changed between prime and probe. Thus, repeated distractors (both to-be-attended and to-be-unattended ones) appeared in each of the possible distractor locations with the same probability on the probe, regardless of their position on the prime display.
Method
Participants
Fifteen students (14 women, one man) from the University of Trier took part in Experiment 1b. Their median age was 20 years, with a range from 18 to 27 years. All participants took part in exchange for partial course credit.
Design
The design of Experiment 1b was the same as in Experiment 1a. In addition, in Experiment 1b, the position of the to-be-attended distractors repeated from prime to probe on half of the trials and changed between prime and probe on the other half.
Materials and procedure
The materials and procedure were the same as in Experiment 1a, with the following exceptions. All participants were instructed to attend to distractors in the upper left and lower right corners on half of the displays and to distractors in the upper right and lower left corners on the other half. The to-be-attended location could change between trials, as well as between the prime and probe within one trial (see Fig. 1b). To-be-attended distractor positions on the prime and probe were indicated before each display. After participants had started a trial by pressing the space bar, in Experiment 1b, a display appeared for 300 ms, indicating at which locations the to-be-attended distractors would appear. This display included a white fixation mark at the center (at the location of the target letter) and two red exclamation marks at the locations of the to-be-attended distractors. Then, the screen went blank for 200 ms before the prime display appeared. Similarly, after the prime response a white fixation mark and two red exclamation marks were shown to indicate the relevant distractor positions on the probe display. After another 200-ms blank screen, the probe display was presented until participants’ responses. Everything else was exactly the same as in Experiment 1a.
Results
Only trials with correct answers to the prime and the probe were considered. Reaction times that were more than 1.5 interquartile ranges above the third quartile of the distribution for the entire sample (Tukey, 1977) or that were shorter than 200 ms were excluded from the analysis. Due to these constraints, 7.8 % of all trials were discarded (probe error rate was 1.9 %, and prime error rate was 1.7 %). Mean reaction times and error rates for the probe displays are depicted in Table 1.
Reaction times
In a 2 (position relation of attended distractor: position change vs. position repetition between prime and probe) × 2 (instructed attention: distractors to be attended vs. unattended) × 2 (response relation: response repeated vs. response changed) × 2 (distractor relation: distractor repeated vs. distractor changed) MANOVA with Pillai’s trace as the criterion, the four-way interaction was not significant, F(1, 14) < 1, p > .5, ηp 2 = .03. That is, any modulation of the Response Relation × Distractor Relation interaction by instructed attention did not differ for trials in which the positions of the to-be-attended distractors changed from prime to probe and those in which the to-be-attended positions were the same on prime and probe.Footnote 2 Therefore, we dropped the factor of Position Relation from all further analyses. In a 2 (instructed attention: distractors to be attended vs. unattended) × 2 (response relation: response repeated vs. response changed) × 2 (distractor relation: distractor repeated vs. distractor changed) MANOVA with Pillai’s trace as the criterion, the main effects of response relation and distractor relation were significant, F(1, 14) = 6.28, p = .025, ηp 2 = .31, for response relation, and F(1, 14) = 6.39, p = .024, ηp 2 = .31, for distractor relation. Responses to repeated targets (M = 1,030 ms, SD = 260 ms) were faster than those to changed targets (M = 1,144 ms, SD = 239 ms), and responses were faster if distractors were repeated (M = 1,067 ms, SD = 228 ms) than if the distractors changed from prime to probe (M = 1,103 ms, SD = 241 ms). Neither the Instructed Attention × Response Relation interaction, F(1, 14) = 3.23, p = .094, ηp 2 = .19, nor the Instructed Attention × Distractor Relation interaction, F(1, 14) = 1.62, p = .224, ηp 2 = .10, was significant. More interestingly, the interaction of response relation and distractor relation was significant, F(1, 14) = 20.79, p < .001, ηp 2 = .60, indicating that distractor repetition led to a larger advantage in response repetition than in response change trials. Most importantly, once again the three-way interaction of instructed attention, response relation, and distractor relation was significant, F(1, 14) = 4.72, p = .048, ηp 2 = .25. Separate analyses revealed a significant interaction of response relation and distractor relation for to-be-attended distractors, F(1, 14) = 50.67, p < .001, ηp 2 = .78, but no interaction of response relation and distractor relation for to-be-unattended distractors, F(1, 14) = 0.40, p = .537, ηp 2 = .03.
Error rates
The same MANOVA on error rates revealed similar results. The main effects of response relation, F(1, 14) = 21.74, p < .001, ηp 2 = .61, and distractor relation, F(1, 14) = 7.26, p = .017, ηp 2 = .341, were significant, as well as the interaction of response relation and distractor relation, F(1, 14) = 6.47, p = .023, ηp 2 = .316. Most importantly, the three-way Response Relation × Distractor Relation × Instructed Attention interaction was also significant, F(1, 14) = 5.36, p = .036, ηp 2 = .28, indicating that the interaction of response relation and distractor relation was significant for to-be-attended distractors, F(1, 14) = 9.73, p = .008, ηp 2 = .41, but not for to-be-unattended distractors, F(1, 14) < 1, p > .8, ηp 2 = .004.
Comparison of Experiments 1a and 1b
In a 2 (experiment: 1a vs. 1b) × 2 (instructed attention: distractors to be attended vs. unattended) × 2 (response relation: response repeated vs. response changed) × 2 (distractor relation: distractor repeated vs. distractor changed) mixed model MANOVA with Pillai’s trace as the criterion, the main effect of experiment, F(1, 38) = 443.94, p < .001, ηp 2 = .92, and the interaction of response relation and experiment were significant, F(1, 37) = 6.78, p = .013, ηp 2 = .16. Importantly, the four-way interaction of instructed attention, response relation, distractor relation, and experiment was not significant, F(1, 37) = 0.33, p = .571, ηp 2 = .009. The effects that instructed attention had on distractor–response binding did not differ between Experiments 1a and 1b. None of the other effects that included the Experiment factor reached significance. In the same MANOVA on error rates, none of the interactions that included the Experiment factor were significant.
Discussion
As in Experiment 1a, we found an interaction of response relation and distractor relation, which can be interpreted as a general effect of distractor–response binding. Moreover, this interaction was also specified by an interaction of instructed attention, response relation, and distractor relation. That is, as in Experiment 1a, the effect of distractor–response binding was modulated by instructed attention to the distractors.
In sum, the results of Experiment 1 clearly evidence that an additional response-irrelevant stimulus is integrated into an event file with the response if participants are instructed to attend to this stimulus. In contrast, to-be-unattended distractors were not integrated into event files. That is, the attention distributed to a stimulus can indeed modulate binding effects of this stimulus with a co-occurring response. Moreover, Experiment 1b evidenced that the effect the to-be-attended distractors had on responses did not hinge on a repetition of distractor location. In fact, the influence instructed attention had on distractor–response binding did not differ for constant and variable relevant positions. This can be taken as an indication that distractor identity retrieved prime responses independent of distractor location.
Since the results indicate that participants indeed attended to the distractors they were instructed to attend to and ignored the other distractors, we can assume that the spatial distribution of attention changed between the prime and the probe response, in half of the trials. Note that whether or not this distribution changed did not modulate the influence instructed attention had on the effect of distractor–response binding. This indicates that spatial distribution of attention at the time of responding does not become part of the event file.
Moreover, the results of Experiment 1 are in line with evidence that association formation between stimuli and responses can be influenced by attention that is guided via a preceding cue (Logan & Etherton, 1994, Exp. 5). In addition to earlier results, the present findings indicate that the attentional distribution affects bindings that are formed during a single encounter of distractor–response pairs.
Taken together, the results of Experiments 1a and 1b indicate that attention to response-irrelevant stimuli modulates integration of these additional stimuli with the responses, but that the location of a repeated distractor does not play a crucial role in the modulation of distractor–response binding. Apparently the distractor identity, independent of the distractor location, is integrated with, and later on can retrieve, the response. With target stimuli always appearing at the center of the screen, cueing the to-be-attended distractors spatially prevented every search of the presented stimulus set. Instead, attention could be adjusted to the relevant locations before stimulus onset. Experiment 2 was intended to investigate whether the modulation of distractor–response binding by attention also holds if attention is guided via relevant stimulus features and the display has to be searched for the to-be-attended distractors.
Experiment 2
Experiment 1 showed that spatially distributing attention to the location that a distractor is about to occupy ensures integration of the stimulus appearing in the attended location with the executed response, even if the stimulus is entirely irrelevant for this response. In Experiment 2, we used the same stimulus arrangement and task as in Experiment 1 and tested whether feature-based attention influences distractor–response binding effects in a similar way.
In many cases (e.g., in situations requiring visual search of a scene), attention is not guided by a visual cue but according to stimulus features. Spatial and feature-based attention can be assumed to differ from each other regarding time courses and mechanisms (e.g., Ling, Liu, & Carrasco, 2009; Liu, Stevens, & Carrasco, 2007). Moreover, attentional shifts between colors at one location seem to be associated with activation in different subpopulations of neurons than attention shifts between locations (Greenberg, Esterman, Wilson, Serences, & Yantis, 2010). Regarding these general distinctions, one might also assume that stimulus–response integration is differently influenced by spatial and feature-based attention. In addition, Logan and Etherton (1994) found less learning of co-occurrences if to-be-attended stimuli were cued than if participants had to search for to-be-attended stimuli. They concluded that less attention distributed to distractors in the cued condition than in the search condition led to fewer instances of integration of the distractors. Regarding these findings, it is not surprising that we found no distractor–response binding for the to-be-unattended distractors in Experiments 1a and 1b. Requiring participants to search for the to-be-attended distractor, however, might lead to a different result.
To test this assumption, participants again identified target letters that were surrounded by distractor stimuli and were instructed to attend to some of the distractors while completely ignoring the others. Importantly, to-be-attended distractors were no longer indicated by position cues, but in Experiment 2 were defined via the stimulus set they were drawn from. Thus, participants were not instructed to attend to a certain space, but to the features identifying certain objects. As in Experiment 1b, the locations of to-be-attended stimuli could change between prime and probe displays. However, participants were never informed of such a location change before stimulus onset, but were forced to scan each display in order to identify the locations (and the identity) of the to-be-attended distractors. The same pattern of results as in Experiments 1a and 1b would indicate that the type of attentional control does not play a crucial role in the modulating effect that attention has on distractor processing.
Method
Participants
A total of 32 students (27 women, five men) from the University of Trier took part in the experiment. Their median age was 21 years, with a range from 18 to 30 years. All participants took part in exchange for partial course credit.
Design, materials, and procedure
The design, materials, and procedure were the same as in Experiment 1b, with the following exceptions. The to-be-attended distractor digits were not defined by location but by identity: Half of the participants were instructed to attend to the digits 1–4 and ignore the digits 5–8, whereas the other half received the opposite mapping. As in Experiment 1b, the locations of the to-be-attended distractors could change both between trials and between prime and probe displays within one trial. However, throughout the experiment none of the locations were cued. The sequence of events in one trial was therefore similar to that in Experiment 1a: Participants started each trial by pressing the space bar. After 500 ms of a blank screen, the prime display appeared and stayed on the screen until participants responded. After the prime response, another blank display of 500-ms duration appeared, before the probe display was presented. As with the prime, the probe target and distractors stayed on the screen until participants responded. As in Experiment 1, on one third of the trials the participants were then prompted to report the identities of the to-be-attended distractor digits in the second-to-last (i.e., the prime) and the last (i.e., the probe) displays. Unlike in the previous experiments, these prompts consisted only of the question “Relevant number on the second-to-last/last display?” and did not include question marks indicating the distractor positions on the prime and probe displays.
Results
According to the same criteria as in Experiment 1b, 7.6 % of all trials were discarded (the probe error rate was 1.9 %, and the prime error rate was 1.7 %). The mean reaction times and error rates for the probe displays are depicted in Table 2.
Reaction times
In a 2 (position relation of attended distractor: position change vs. position repetition between prime and probe) × 2 (instructed attention: distractor to be attended vs. unattended) × 2 (response relation: response repeated vs. response changed) × 2 (distractor relation: distractor repeated vs. distractor changed) MANOVA with Pillai’s trace as the criterion, the four-way interaction was not significant, F(1, 31) < 1, p > .3, ηp 2 = .03. That is, any modulation of the Response Relation × Distractor Relation interaction by instructed attention was not different for trials in which the positions of the to-be-attended distractors changed from prime to probe and those in which the to-be-attended positions were the same from prime to probe.Footnote 3 Therefore, we dropped the factor of Position Relation from all further analyses. In a 2 (instructed attention: distractor to be attended vs. unattended) × 2 (response relation: response repeated vs. response changed) × 2 (distractor relation: distractor repeated vs. distractor changed) MANOVA with Pillai’s trace as the criterion, the main effects of response relation and instructed attention were significant: F(1, 31) = 140.87, p < .001, ηp 2 = .82, for response relation, and F(1, 31) = 8.97, p = .005, ηp 2 = .22, for instructed attention. Responses to repeated targets (M = 795 ms, SD = 182 ms) were faster than those to changed targets (M = 1,073 ms, SD = 261 ms), and responses were faster if the (potentially) repeated distractors were to be attended (M = 918 ms, SD = 208 ms) than if the repeated distractors were to be unattended (M = 937 ms, SD = 215 ms). Neither the Instructed Attention × Response Relation interaction, F(1, 31) = 4.09, p = .052, ηp 2 = .12, nor the Instructed Attention × Distractor Relation interaction, F(1, 31) = 1.57, p = .219, ηp 2 = .05, reached significance. The interaction of response relation and distractor relation was also not significant, F(1, 31) = 3.36, p = .078, ηp 2 = .10. Most importantly, again the three-way interaction of instructed attention, response relation, and distractor relation was significant, F(1, 31) = 13.46, p = .001, ηp 2 = .30. That is, the interaction of response relation and distractor relation was modulated by instructed attention to the distractors. Separate analyses revealed a significant interaction of response relation and distractor relation for to-be-attended distractors, F(1, 31) = 10.66, p = .003, ηp 2 = .26, but no interaction of response relation and distractor relation for to-be-unattended distractors, F(1, 31) = 1.01, p = .323, ηp 2 = .03.
Error rates
The same MANOVA on error rates revealed similar findings. The main effect of response relation, F(1, 31) = 58.20, p < .001, ηp 2 = .65, and the interaction of response relation and distractor relation, F(1, 31) = 4.77, p = .037, ηp 2 = .13, were significant. The three-way Response Relation × Distractor Relation × Instructed Attention interaction was not significant, F(1, 31) = 3.28, p = .080, ηp 2 = .10. Yet, in separate analyses, the interaction of response relation and distractor relation was significant for to-be-attended distractors, F(1, 31) = 6.98, p = .013, ηp 2 = .18, but not for to-be-unattended distractors, F(1, 31) < 1, p > .6, ηp 2 = .009.
Discussion
As in Experiment 1, the interaction of instructed attention, response relation, and distractor relation in Experiment 2 indicated an effect of distractor–response binding for to-be-attended distractors, whereas we found no evidence for distractor–response binding if stimuli were to be unattended. That is, a spatial focus of attention on the location of a response-irrelevant stimulus before stimulus onset was not a prerequisite for binding between this stimulus and the executed response. Instead, feature-based attention seemed to have the same effect on response integration and retrieval as had the spatial distribution of attention. That is, even if attention was deployed to a stimulus after stimulus onset according to the features of the current stimuli, the integration of response-irrelevant stimuli and responses was modulated by whether or not attention was focused on a stimulus at the time of responding. More attention appeared to be necessary for an immediate consequence of a single instance than for a similar effect after several instance repetitions (see Logan & Etherton, 1994). However, as with associations formed by repeated pairings of distractors and responses, attention in our Experiment 2 again played a role in the effects of single distractor–response combinations. Together, Experiments 1a, 1b, and 2 might be a first indication that stimulus–response bindings are the immediate effects of instance integration, as was proposed in the instance theory of automatization.
Note that according to the feature integration theory (Treisman & Gelade, 1980), it is necessary to direct attention to the location of an object in order to integrate the perceived features into one object file. In this regard, we can assume that even the rejected distractor digits received some attention that ensured object integration. However, this kind of attention was not sufficient for the distractor to be integrated with, and later on retrieve, the response to the target.
In all of Experiments 1a, 1b, and 2, participants were instructed either to attend to or not to attend to repeated distractors on both the prime and the probe displays. Therefore, none of the experiments provided evidence for whether attention modulates the binding of distractors and responses, the retrieval of the responses, or both processes. In a third experiment, we aimed to pinpoint the particular process that is influenced by attention being distributed to the distractor stimulus.
Experiment 3
Effects of stimulus–response binding are typically evidenced by the responses that repeated stimuli retrieve: Participants are faster and/or more accurate at responding if the retrieved response matches the required response than if the retrieved response differs from the required response. If this pattern is found, the assumption is that the retrieved response was bound to the presented stimulus at an earlier point in time. Thus, retrieval theories generally assume at least two separate processes—namely, stimulus–response integration and stimulus–response retrieval. Hence, additional factors, such as the focus of attention, may separately influence integration and retrieval. Experiment 3 was designed to investigate which of these processes is influenced by instructed attention. It has been shown that priming effects that can be interpreted as evidence for retrieval processes are influenced by the role that the repeated stimulus has at response retrieval (i.e., on the probe), but not by its role at stimulus–response binding (i.e., on the prime; Ihrke, Behrendt, Schrobsdorff, Herrmann, & Hasselhorn, 2011). Regardless of whether a repeated stimulus was the target or the distractor at encoding (i.e., on the prime), the effects of response retrieval were larger if the repeated stimulus was the probe target than if it was the probe distractor. In addition, it has been speculated that the effects of stimulus–response binding depend on the attentional distribution at retrieval rather than on that at binding (e.g., Hommel, Memelink, Zmigrod, & Colzato, 2013; Zmigrod, Spapé, & Hommel, 2009). With these findings in mind, it can be guessed that the attentional distribution at the time of response retrieval, but not at the time of stimulus–response integration, influenced the effect of distractor–response binding in Experiments 1 and 2. Yet, since a to-be-attended distractor was always to be attended on the prime and the probe, and a to-be-unattended distractor was not instructed to be attended on either the prime or the probe, it was not possible to pinpoint the process that was influenced by attentional distribution. To disentangle whether the influence of attention works on the integration, the retrieval, or both processes of distractor–response binding, the likelihood of attention being distributed to the (potentially) repeated distractor on the prime and the probe was varied orthogonally. That is, participants attended to certain locations throughout the experiment, and repeated distractors could appear in those positions on either the prime or the probe. If attention to the distractor is necessary for the integration of this stimulus with the currently executed response, but not for the retrieval of this response, the effect of distractor–response binding would be revealed for to-be-attended prime distractors, independent of whether or not these distractors were again to be attended on the probe. If binding also occurs without attention to the integrated stimuli, but attention to the distractors is essential to trigger response retrieval, we would find an effect of distractor–response binding for distractors that were to be attended on the probe, independent of the instructed distribution of attention on the prime. Finally, if attention to the distractors is necessary both for binding and for retrieval to take place, we would find evidence for distractor–response binding only for trials in which the repeated distractor was to be attended on both the prime and the probe.
Method
Participants
A total of 29 students (26 women, three men) from the University of Trier took part in the experiment. Their median age was 21 years, with a range from 18 to 28 years. All participants took part in exchange for partial course credit.
Design
The design essentially comprised four within-subjects factors—namely, Response Relation (repetition vs. change), Distractor Relation (repetition vs. change), Instructed Attention to the Prime Distractors (distractors to be attended vs. unattended), and Instructed Attention to the Probe Distractors (distractors to be attended vs. unattended).
Materials and procedure
The materials and procedure were the same as in Experiment 1a, with the following exceptions. Each of the eight possible distractor digits was sometimes to be attended and sometimes to be unattended. As in Experiment 1a, half of the participants were instructed to attend to the upper left and lower right distractor positions and to ignore the upper right and lower left distractor positions, whereas the other half received the opposite mapping. In addition to the to-be-attended (i.e., the repeated distractor appears in a to-be-attended location on both the prime and the probe) and to-be-unattended (i.e., the repeated distractor appears in a to-be-unattended location on both the prime and the probe) conditions of Experiment 1a, the repeated distractor identity could also appear in a to-be-attended position on the prime and a to-be-unattended position on the probe, and it could appear in a to-be-unattended position on the prime and a to-be-attended position on the probe. Thus, each of the four conditions (response repetition/distractor repetition, response repetition/distractor change, response change/distractor repetition, and response change/distractor change) was realized 12 times in each of the four conditions resulting from instructed attention (repeated distractor to be attended vs. unattended) being orthogonally varied on the prime and the probe: to be attended on the prime–to be attended on the probe, to be attended on the prime–to be unattended on the probe, to be unattended on the prime–to be attended on the probe, and to be unattended on the prime–to be unattended on the probe. The identity of the distractors in the other positions, respectively, changed from prime to probe.
Results
According to the same criteria as in Experiment 1a, 7.4 % of all trials were discarded (the probe error rate was 1.7 %, and the prime error rate was 1.8 %). Mean reaction times and error rates for probe displays are depicted in Table 3.
Reaction times
In a 2 (instructed prime attention: distractor to be attended vs. unattended) × 2 (instructed probe attention: distractor to be attended vs. unattended) × 2 (response relation: response repeated vs. response changed) × 2 (distractor relation: distractor repeated vs. distractor changed) MANOVA with Pillai’s trace as the criterion, the main effect of response relation, F(1, 28) = 68.62, p < .001, ηp 2 = .71, and the interaction of instructed prime attention and instructed probe attention, F(1, 28) = 12.24, p = .002, ηp 2 = .30, were significant. Responses to repeated targets (M = 755 ms, SD = 307 ms) were faster than those to changed targets (M = 996 ms, SD = 369 ms); also, if the potentially repeated distractor was to be attended on the probe, responses were faster if this distractor had also been a to-be-attended one on the prime than if it had been a to-be-unattended one on the prime, whereas instructed attention on the prime did not make a difference if the repeated distractor was to be unattended on the probe. Neither the interaction of instructed prime attention and distractor relation, F(1, 28) = 4.06, p = .054, ηp 2 = .13, nor the interaction of instructed probe attention and distractor relation, F(1, 28) < 1, p > .8, ηp 2 = .001, was significant. More importantly, the interaction of response relation and distractor relation was significant, F(1, 28) = 20.71, p < .001, ηp 2 = .43, as well as the three-way interaction of instructed probe attention, response relation, and distractor relation, F(1, 28) = 5.14, p = .031, ηp 2 = .16. Separate analyses revealed a significant interaction of response relation and distractor relation if distractors were to be attended on the probe, F(1, 28) = 16.32, p < .001, ηp 2 = .37, but not if distractors were to be unattended on the probe, F(1, 28) = 1.80, p = .190, ηp 2 = .06. Neither the three-way interaction of instructed prime attention, response relation, and distractor relation, F(1, 28) = 1.79, p = .192, ηp 2 = .06, nor the four-way interaction of instructed prime attention, instructed probe attention, response relation, and distractor relation, F(1, 28) = 2.66, p = .114, ηp 2 = .09, was significant. For the sake of completeness, the Instructed Prime Attention × Instructed Probe Attention × Response Relation interaction, F(1, 28) = 4.24, p = .049, ηp 2 = .13, was significant, as well.
Error rates
The same MANOVA on error rates revealed a similar pattern. The main effects of response relation, F(1, 28) = 27.78, p < .001, ηp 2 = .50, and instructed probe attention, F(1, 28) = 4.33, p = .047, ηp 2 = .13, were significant, as was the Instructed Prime Attention × Response Relation interaction, F(1, 28) = 13.53, p = .001, ηp 2 = .33. The main effect of instructed prime attention, F(1, 28) = 3.77, p = .062, ηp 2 = .12; the Response Relation × Distractor Relation interaction, F(1, 28) = 3.91, p = .058, ηp 2 = .12; the three-way Instructed Probe Attention × Response Relation × Distractor Relation interaction, F(1, 28) = 2.94, p = .098, ηp 2 = .10; the three-way Instructed Prime Attention × Response Relation × Distractor Relation interaction, F(1, 28) < 1, p > .4, ηp 2 = .02; and the four-way Instructed Prime Attention × Instructed Probe Attention × Response Relation × Distractor Relation interaction, F(1, 28) < 1, p > .6, ηp 2 = .006, were not significant.
Discussion
The interaction of instructed probe attention, response relation, and distractor relation indicated that repeated distractors triggered response retrieval with a higher probability if distractors were to be attended on the probe than if distractors were to be unattended on the probe. This interaction was not further modulated by the attention modulation on the prime display. That is, instructed attention at integration of the distractor and response did not have a large influence on whether or not response retrieval was eventually triggered. Put differently, distractor–response retrieval only occurred if participants were instructed to focus attention on the stimulus triggering retrieval, but not if the potentially retrieving stimulus was to be unattended. In contrast, distractor–response binding seemed to be sufficient to enable later retrieval processes, independent of whether or not attention had been distributed to the stimulus at integration. It should be noted that all of the presented distractor identities (i.e., also the identities of the to-be-unattended distractors) in Experiment 3 were relevant on approximately half of the trials throughout the experiment. Therefore, it is likely that even the to-be-unattended distractors captured some attention. Apparently, this kind of attention was sufficient for the stimuli to be integrated with the executed response, whereas it was not sufficient for the stimulus to start a retrieval process.
This pattern is in line with findings by Ihrke et al. (2011), who showed that priming effects were modulated by the role of the repeated stimulus on the probe, but not by its role on the prime. The present results also indicate that an influence of attentional resources during stimulus encoding on response retrieval effects cannot be large. This is also in line with Hommel’s (2005; Hommel & Colzato, 2004) assumption that hardly any attention is necessary for the integration of a stimulus into an event file. However, at first sight these results seem to be contrary to several studies that have shown attention to be at least as important during encoding as during retrieval in an automatization process (e.g., Boronat & Logan, 1997; Logan, & Etherton, 1994). However, Logan and colleagues investigated S–R retrieval that had been learned after several repetitions of a particular stimulus–response pairing, whereas we were concerned with retrieval of a stimulus–response pairings after a single encounter. In addition, the retention intervals were quite different between the studies; they could include several trials, rest phases, and sometimes instructions in the studies of Logan and colleagues, whereas the retention interval in our experiment was less than 2 s. We will come back to these differences in the General Discussion.
General discussion
The aim of the present study was to investigate whether attention needs to be deployed to a response-irrelevant stimulus for it to be integrated with and subsequently retrieve a response. We used stimuli that were entirely response-irrelevant and instructed participants to attend only to some of these additional stimuli, while ignoring the others. Repeated additional stimuli retrieved responses if they were to be attended, but not if they were to be unattended (Exp. 1a). Furthermore, the influence of instructed attention on the binding and retrieval of additional stimuli seems to be independent of whether attention is spatially controlled (Exp. 1b) or feature-based (Exp. 2). Finally, the modulation of the distractor–response binding effect via instructed attention to distractors was regulated by the instructed attentional distribution at response retrieval, whereas the instructed attentional distribution at the integration of stimulus and response did not seem to have a large influence on the effect (Exp. 3). For an overview of the results, see Fig. 2.

Distractor–response binding effects as a function of instructed attention to the distractors on the probe (distractor to be attended vs. unattended) in all experiments. Distractor–response binding effects were calculated as the difference between distractor repetition effects in response repetition and response change trials. Note that to-be-attended (or to-be-unattended) probe distractors were also to be attended (or to be unattended) on the prime in Experiments 1a, 1b, and 2. In Experiment 3, for both to-be-attended and to-be-unattended probe distractors, prime distractors were to be attended on half of the trials and to be unattended on the other half. D, distractor
Notably, the way that attention was manipulated did not play an important role for the reported modulation in the present experiments. Instructed attention to response-irrelevant stimuli had the same effect on stimulus–response integration and retrieval when it was guided by stimulus features as when it was adjusted via a spatial cue. Although spatial and feature-based attentional processes have been suggested to differ (e.g., Ling et al., 2009; Liu et al., 2007), stimulus–response binding processes seem to be modulated solely by whether or not attention is focused on the stimulus at the time of responding. Interestingly, this was the case even though the participants in Experiments 1 and 3 were able to spatially adjust their attentional focus before stimulus onset, whereas attentional adjustment via object features in Experiment 2 was only possible after target and distractor onset. That is, even though relevant stimulus features (such as orientation and angle of lines) were attended in both distractor sets in Experiment 2, only to-be-attended distractors were integrated with and could retrieve a response. In fact, the present findings might even indicate that feature-based and location-based attention do not differ so much, after all.Footnote 4 This would also be in line with the suggestion that both kinds of attentional control are guided by the same functional system (Yantis & Serences, 2003). On the basis of the present results, we cannot decide whether or not attention differs depending on the way it is controlled. Yet, the kind of attentional control (i.e., spatial vs. feature-based attention) seems to be irrelevant for the role that attention plays in stimulus–response binding.
At first glance, our results regarding the to-be-unattended distractors seem to be inconsistent with recent findings by Giesen et al. (2012). In all of the present experiments, the interaction of response relation and distractor relation (i.e., the effect of distractor–response binding) did not reach significance if the distractors were to be unattended. Even when we combined the data from all four experiments, this effect was not significant [F(1, 99) = 0.08, p = .772, ηp 2 = .001; the power was at 1 – β = .80]. Thus, one can conclude that at least a large, or even medium-sized, effect of distractor-based response retrieval is unlikely if distractors are unattended. In contrast to our findings for the to-be-unattended distractors, Giesen et al. found significant effects of distractor–response binding for distractors that were not particularly attended. The main differences between the present experiments and the aforementioned study were the number of presented distractors (two vs. four) and the manner of distractor arrangement. Neill, Valdes, Terry, and Gorfein (1992) pointed out that with more distractors, the probabilities are reduced for both the encoding of all distractors and the episodic retrieval of the priming episode (for less negative priming with a larger number of distractors, see Neumann & DeSchepper, 1992). Thus, the number of distractors may have been a factor that contributed to ensure encoding and retrieval processes, regardless of any instruction in the study by Giesen and colleagues, whereas this factor reduced the probabilities of encoding and retrieval processes in the present setup. With the four distractors in the present experiments, the saliency of each distractor set was reduced, and a smaller portion of the display was repeated (two out of four, instead of two out of two distractors). Therefore, a probe display including two repeated and two changed distractor identities did not serve as a good trigger for retrieval, unless participants intentionally allocated attention to the repeated distractors. In addition, the arrangement of distractor stimuli may have contributed to the difference in the results. Whereas Giesen and colleagues presented target and distractors in a horizontal line, the distractors in the present study were diagonally aligned to the targets. It has been shown that distractor–response binding is possible with the former stimulus alignment, yet no evidence exists regarding the latter arrangement. Yet it can be assumed that grouping principles regarding target and distractor stimuli modulate binding processes (Frings & Rothermund, 2011). Therefore, it is possible that the stimulus configuration in the present experiments prevented distractor–response binding of the to-be-unattended distractors.
In line with the results of Ihrke et al. (2011), the present Experiment 3 provided evidence that attentional modulation of the distractor–response binding effect is largely based on differences in retrieval processes. The effect of distractor–response binding reached significance only in trials that required participants to distribute focused attention to the distractor that triggered retrieval, regardless of whether or not participants had also been instructed to attended to this stimulus during the prime. Similarly, Ihrke and colleagues investigated encoding and retrieval mechanisms in a negative-priming paradigm and manipulated whether a repeated stimulus was presented as the target or as the distractor (note that targets typically receive more attention than distractors). They found retrieval processes to be influenced by the task relevance of the repeated stimulus: Repetition effects were larger if the repeated stimulus was presented as the probe target than if it was presented as the probe distractor. In addition to the findings by Ihrke and colleagues, our results indicate that response relevance is not necessary for a modulation of response retrieval. Distributing attention to the repeated stimulus seems to be sufficient to influence retrieval processes.
It is noteworthy that the distractor–response binding effect was not significantly modulated by whether or not a distractor was to be attended on the prime (i.e., at the time the binding took place). Thus, binding of distractors and responses occurred even for stimuli that did not receive focused attention. These findings are in line with Hommel and Colzato’s (2004) suggestion that the determining factor for the integration of stimulus features into an event file is not attention to the feature, but whether or not this feature varies on a dimension that is relevant in the present situation. As we mentioned above, our results also indicate that this sort of relevance is not sufficient for the retrieval of a response due to the repetition of a stimulus.
However, it should be noted that the four-way interaction of instructed prime attention, instructed probe attention, response relation, and distractor relation was not far from being significant (p = .114, ηp 2 = .09). In addition, the achieved power for this test was not very large (1 – β = .5). Therefore, we cannot exclude the possibility that instructed attention during the prime would also influence the results if considerably larger sample sizes were used. This would be similar to studies by Logan and colleagues (e.g., Boronat & Logan, 1997; Logan & Etherton, 1994; Logan et al., 1996, 1999) indicating that attention at encoding influenced the formation of stimulus–response associations.
Finally, an interesting point of the present results is that in contrast to the attentional modulation of distractor–response binding effects, the effect of distractor inhibition (indicated by the main effect of distractor relation) was not far from being modulated by attention to the prime distractor [F(1, 28) = 4.06, p = .054, ηp 2 = .13], but was not modulated by attention to the probe distractor [F(1, 28) < 1, p > .8, ηp 2 = .001]. These results regarding distractor repetition effects are highly plausible. The inhibition account (see Frings, Wentura, & Wühr, 2012; Frings & Wühr, 2007; Houghton & Tipper, 1994) predicts that distractor inhibition is not influenced by mechanisms on the probe, because it is caused by a residual deactivation of the internal representation of the prime distractor. Yet, processing resources and the focus of attention on the prime influence the degree to which a distractor is processed and inhibited in order to ensure fast and correct responses on the prime. This finding also adds to the evidence that distractor–response binding and distractor inhibition are two independent processes in action control (see Giesen et al., 2012), evidencing that the two processes are modulated by the attentional distributions at different points in time in a prime–probe sequence.
Implications of the present results
The present findings are interesting not only with regard to the binding literature, but may have important implications in several different fields. Regarding the aforementioned research by Logan and colleagues, one may ask if the present results are in fact an indication of event file integration and retrieval, or if they can also be interpreted in terms of association formation. First, we need to point out that the process of binding is assumed to lead to a number of short-lived binary conjunctions, which can be thought of as a loose network of clusters rather than as one master file (Hommel, 1998, 2004). Thus, interpreting the present effects as being due to event file retrieval implicitly assumes the existence of short-lived associations between the responses and distractors. A more interesting question is whether the binding process that leads to such temporary conjunctions also plays a role in association learning (i.e., the formation of longer-lasting associations). In terms of the instance theory of automatization (Logan, 1988, 1990), this possibility seems realistic. Notably, studies investigating automatization (e.g., Boronat & Logan, 1997; Logan & Etherton, 1994; Logan et al., 1996, 1999) have used comparatively long retention intervals (including several trials, rest phases, and sometimes instructions). It can be assumed that these intervals led to retrieval from long-term memory in the transfer phases. In contrast, with prime–probe intervals of less than 2 s, the present retrieval processes took place most likely within working memory. Hence, the observed effects can be understood as immediate consequences of single instance encodings. In turn, the present results, as well as those of other studies regarding binding processes, may add to the understanding of the process of association learning.
This speculation also suggests a different interpretation of the aforementioned studies on association learning. None of the experiments was designed to test the immediate consequences of instance integration separately. Moreover, during the encoding phases in these studies, stimulus–response pairs were presented repeatedly (e.g., Boronat & Logan, 1997; Logan et al., 1996, 1999). Therefore, all but the first presentation of a given stimulus during encoding would be assumed to trigger automatic retrieval of the earlier instances. Hence, retrieval (as analyzed in the present experiments) may have taken place during encoding and could have been influenced by the attentional distribution during this phase. In contrast, attention during the sort of integration that we investigated was not separately manipulated in those studies. Assuming that we were indeed investigating single instances, the present results suggest that attention at the repeated encodings increased automatic retrieval already during the encoding phases. Thus, it might be speculated that repeated retrieval of former co-occurrences plays a role in the formation of long-term associations. This may also be the reason why Logan and colleagues found significant influences of attention during encoding, whereas our results indicated no large modulation of stimulus–response integration due to attention.
Regarding findings of rapid stimulus–response learning in long-term repetition-priming paradigms (e.g., Dennis & Schmidt, 2003; Dobbins, Schnyer, Verfaellie, & Schacter, 2004; Horner & Henson, 2009, 2011), one might speculate that the investigated bindings could also more directly lead to learning. For example, similar to the present experiments, Horner and Henson (2009, 2011) investigated retrieval effects after a single encounter of stimuli and found retrieval of response codes (as well as retrieval of decision and classification codes) at stimulus repetition. Importantly, these effects occurred with 10- to 15-min retention between prime encoding and probe response, indicating single-trial learning of stimulus–response associations. An important difference between these studies and the present paradigm is the role of the repeated stimuli. The mentioned long-lasting retrieval effects were found for target-to-target repetitions, whereas we investigated retrieval due to distractor repetitions. In addition, Horner and Henson (2009, 2011) used a large stimulus set, and participants saw each target maximally twice throughout an experiment. With the limited stimulus set that we used, distractors were frequently repeated with each of the possible responses. Hence, we can be relatively certain that distractor–response associations in the present experiments were not as long-lasting. Moreover, it generally seems unlikely that after a single encounter, distractor–response associations can reach a strength similar to that of target–response associations. What exactly it takes for a distractor stimulus to become part of such rapid learning is an important question for future research.
However, on the basis of the present findings, we can speculate that the representations of all presented stimuli are generally activated to some extent, and that attention can boost such activation. If activation then decays over time, it is reasonable to assume that representations of formerly attended (more strongly activated) stimuli have an activation advantage for longer time periods than do ignored items. In addition, formerly attended features might have an advantage due to bindings or associations to other attended features: Activation in one feature might reinforce activation in the associated features, generally prolonging activation advantagesFootnote 5.
In contrast to our present findings, studies investigating episodic memory have typically found a larger influence of attention during encoding than during retrieval (e.g., Craik, Govoni, Naveh-Benjamin, & Anderson, 1996; Tulving, Kapur, Craik, Moscovitch, & Houle, 1994). However, these studies were mainly concerned with the investigation of intentional storage and retrieval of episodes. Encoding is assumed to result in long-term memory storage, and retrieval typically refers to an intentional process of remembering an earlier episode, which is often tested explicitly. In contrast, the retrieval processes in our experiments are assumed to be automatic, and their impact upon behavior was measured implicitly. These differences lead to two important distinctions between episodic memory studies and the present experiments. On the one hand, the retention intervals between encoding and retrieval differ dramatically. The term memory retrieval is mostly used if more than a few seconds pass between encoding and retrieval. This was not the case in the present study, and it is highly unlikely that a long-term memory entry would result from the temporary binding between stimuli and responses. It is conceivable that the retrieval of episodes encoded into long-term memory and the retrieval of episodes upheld in working memory can be influenced differently by attention. On the other hand, the aforementioned studies on episodic memory measured intentional retrieval via explicit tests, whereas we measured unintentional retrieval in a task that could be dubbed implicit. Interestingly, attention that had no effect on association learning during an explicit retrieval test was also found to modulate implicitly tested retrieval processes (Boronat & Logan, 1997; Logan & Etherton, 1994; Logan et al., 1999). Since processing seems to differ between intentional and unintentional retrieval (Rugg, Fletcher, Frith, Frackowiak, & Dolan 1997), it is quite possible that these processes are influenced differently by attention.
The present results might also be interpreted in terms of short-term memory storage (Allen, Baddeley, & Hitch, 2006; Baddeley, 1998; Baddeley, Papagno, & Vallar, 1988). The working memory model of Baddeley and colleagues assumes that feature-binding processes are one function of the episodic buffer. A rather small adjustment of the theory might allow bindings that include action as well as perceptual features. In previous experiments, a concurrent task (counting backward in single steps and steps of three and recall of digit strings) did not hamper the binding of stimulus features (Allen et al., 2006). These findings are in line with the present findings regarding attentional influence during the binding of distractors and responses (i.e., during the prime): Attention does not significantly influence the process of feature integration. Whether or not the binding processes discussed by Allen and colleagues are indeed the same as those in the discussion of event file formation is an interesting question for future research that might have important implications for both traditions.
With the two parallel tasks of responding to the target letters and remembering one of the distractor sets, we also need to consider whether the requirements of the attention-directing task influenced reaction times in the letter identification task. Remembering the same digit for the prime and the probe displays is obviously less demanding on working memory resources than is remembering two different letters. As a consequence, participants can be expected to respond faster if the to-be-attended (and to-be-remembered) distractor repeats from prime to probe. Yet, this facilitation should be identical for response repetition and response change trials. The difference of distractor repetition effects between response repetition and response change trials that was observed in the present experiments cannot be accounted for by dual-task interference.
Finally, since the distractor stimuli were not part of the response set, they did not compete with the target for a response during the target identification task. Therefore, one could argue that they should be considered context information rather than distractors. Hence, the present results may indicate that a repeated context can trigger retrieval of associated responses. This is in line with previous studies, showing that context similarity between prime and probe displays in a negative priming task facilitated retrieval effects (Fox & de Fockert, 1998; Neill, 1997; Wong, 2000). In addition to findings in these studies, our results seem to suggest that the context itself can retrieve prime information if it is sufficiently salient (i.e., attended).
Conclusion
In summary, the results of the presented experiments allow insight into how attention affects the influence of stimulus–response binding on action control. In fact, it seems that the integration of stimuli (even irrelevant ones) and responses can take place without much attention—a finding matching previous assumptions. However, to affect behavior, and possibly also to create stimulus–response routines, temporarily associated responses have to be retrieved by repeated stimulus features. Here, attention plays a more important role. In particular, our data showed that attention to a stimulus modulates whether or not it triggers retrieval, and thereby controls whether stimulus–response bindings can affect behavior. Interestingly, the type of attention (spatial vs. feature-based) seems to be irrelevant—as long as stimuli get some kind of attention, retrieval processes will be boosted.
Notes
Note that since response repetition always included target repetition, bindings between distractors and targets may also have contributed to the present results (see Giesen & Rothermund, 2013).
The main effects of response relation, F(1, 14) = 6.19, p = .026, ηp 2 = .31 and distractor relation F(1, 14) = 6.10, p = .027, ηp 2 = .30, and the interactions of position change and response relation, F(1, 14) = 16.06, p = .001, ηp 2 = .53 (with more facilitation due to response repetition if the position repeated than if the position changed); of response relation and distractor relation, F(1, 14) = 21.54, p < .001, ηp 2 = .61; and of instructed attention, response relation, and distractor relation, F(1, 14) = 5.03, p = .042, ηp 2 = .26, were significant. The interactions of position change and instructed attention, F(1, 14) = 3.321, p = .090, ηp 2 = .19; of position change and distractor relation, F(1, 14) = 4.26, p = .058, ηp 2 = .23; and of position change, response relation, and distractor relation, F(1, 14) = 4.15, p = .061, ηp 2 = .23, did not reach significance. Neither did any of the other effects.
The main effects of position change, F(1, 31) = 25.59, p < .001, ηp 2 = .45 (with faster responses if distractor positions were repeated than if they changed); of instructed attention, F(1, 31) = 9.33, p = .005, ηp 2 = .23; and of response relation, F(1, 31) = 140.45, p < .001, ηp 2 = .82, as well as the interaction of instructed attention, response relation, and distractor relation, F(1, 31) = 12.50, p = .001, ηp 2 = .29, were significant. The interactions of position change and instructed attention, F(1, 31) = 3.38, p = .075, ηp 2 = .10; of position change and response relation, F(1, 31) = 3.01, p = .093, ηp 2 = .09; of instructed attention and response relation, F(1, 31) = 3.79, p = .061, ηp 2 = .11; and of response relation and distractor relation, F(1, 31) = 3.26, p = .081, ηp 2 = .10, did not reach significance. Neither did any of the other effects.
We thank Gordon Logan for pointing out these considerations.
We thank Gordon Logan for pointing out these considerations.
References
Allen, R., Baddeley, A., & Hitch, G. (2006). Is the binding of visual features in working memory resource-demanding? J Exp Psychol Gen, 135, 298–313.
Baayen, R. H., Davidson, D. J., & Bates, D. M. (2008). Mixed-effects modeling with crossed random effects for subjects and items. J Mem Lang, 59, 390–412. doi:10.1016/j.jml.2007.12.005
Baddeley, A. (1998). Recent developments in working memory. Curr Opin Neurobiol, 8, 234–238.
Baddeley, A., Papagno, C., & Vallar, G. (1988). When long-term learning depends on short-term storage. J Mem Lang, 27, 586–595.
Bates, D., Maechler, M., Bolker, B., & Walker, S. (2013). lme4: Linear mixed-effects models using Eigen and S4 (R package version 1.0-5). Retrieved from http://CRAN.R-project.org/package=lme4
Baylis, G., & Driver, J. (1992). Visual parsing and response competition: The effect of grouping factors. Percept Psychophys, 51, 145–162. doi:10.3758/BF03212239
Baylis, G. C., & Driver, J. (1993). Visual attention and objects: Evidence for hierarchical coding of location. J Exp Psychol Hum Percept Perform, 19, 451–470. doi:10.1037/0096-1523.19.3.451
Boronat, C., & Logan, G. D. (1997). The role of attention in automatization: Does attention operate at encoding, or retrieval, or both? Mem Cogn, 25, 36–46.
Craik, F. I. M., Govoni, R., Naveh-Benjamin, M., & Anderson, N. D. (1996). The effects of divided attention on encoding and retrieval processes in human memory. J Exp Psychol Gen, 125, 159–180. doi:10.1037/0096-3445.125.2.159
Dennis, I., & Schmidt, K. (2003). Associative processes in repetition priming. J Exp Psychol Learn Mem Cogn, 29, 532–538.
Dixon, P. (2008). Models of accuracy in repeated-measures designs. J Mem Lang, 59, 447–456.
Dobbins, I. G., Schnyer, D. M., Verfaellie, M., & Schacter, D. L. (2004). Cortical activity reductions during repetition priming can result from rapid response learning. Nature, 428, 317–319. doi:10.1038/nature02400
Duncan, J. (1984). Selective attention and the organization of visual information. J Exp Psychol Gen, 113, 501–517. doi:10.1037/0096-3445.113.4.501
Engel, A., & Singer, W. (2001). Temporal binding and the neural correlates of sensory awareness. Trends Cogn Sci, 5, 16–25.
Fox, E., & de Fockert, J. W. (1998). Negative priming depends on prime–probe similarity: Evidence for episodic retrieval. Psychon Bull Rev, 5, 107–113.
Frings, C. (2011). On the decay of distractor–response episodes. Exp Psychol, 58, 125–131.
Frings, C., & Moeller, B. (2010). Binding targets’ responses to distractors’ locations: Distractor response bindings in a location-priming task. Attention, Perception, & Psychophysics, 72, 2176–2183. doi:10.3758/BF03196693
Frings, C., & Moeller, B. (2012). The horserace between distractors and targets: Retrieval-based probe responding depends on distractor–target asynchrony. Journal of Cognitive Psychology, 24, 582–590.
Frings, C., Moeller, B., & Rothermund, K. (2013). Retrieval of event files can be conceptually mediated. Attention, Perception, & Psychophysics, 75, 700–709. doi:10.3758/s13414-013-0431-3
Frings, C., & Rothermund, K. (2011). To be, or not to be…included in an event file: When are distractors integrated into S–R episodes and used for response retrieval? J Exp Psychol Learn Mem Cogn, 37, 1209–1227.
Frings, C., Rothermund, K., & Wentura, D. (2007). Distractor repetitions retrieve previous responses to targets. Q J Exp Psychol, 60, 1367–1377.
Frings, C., Wentura, D., & Wühr, P. (2012). On the fate of distractor representations. J Exp Psychol Hum Percept Perform, 38, 570–575.
Frings, C., & Wühr, P. (2007). On distractor repetition benefits in the negative-priming paradigm. Vis Cogn, 15, 166–178.
Giesen, C., Frings, C., & Rothermund, K. (2012). Differences in the strength of inhibition do not affect distractor–response bindings. Mem Cogn, 40, 373–387. doi:10.3758/s13421-011-0157-1
Giesen, C., & Rothermund, K. (2011). Affective matching moderates S–R binding. Cognition and Emotion, 25, 342–350. doi:10.1080/02699931.2010.482765
Giesen, C., & Rothermund, K. (2013). Distractor repetitions retrieve previous responses and previous targets: Experimental dissociations of distractor–response and distractor–target bindings. J Exp Psychol Learn Mem Cogn. doi:10.1037/a0035278
Greenberg, A. S., Esterman, M., Wilson, D., Serences, J. T., & Yantis, S. (2010). Control of spatial and feature-based attention in frontoparietal cortex. J Neurosci, 30, 14330–14339. doi:10.1523/JNEUROSCI.4248-09.2010
Hommel, B. (1998). Event files: Evidence for automatic integration of stimulus–response episodes. Vis Cogn, 5, 183–216. doi:10.1080/713756773
Hommel, B. (2004). Event files: Feature binding in and across perception and action. Trends Cogn Sci, 8, 494–500. doi:10.1016/j.tics.2004.08.007
Hommel, B. (2005). How much attention does an event file need? J Exp Psychol Hum Percept Perform, 31, 1067–1082. doi:10.1037/0096-1523.31.5.1067
Hommel, B. (2007). Feature integration across perception and action: Event files affect response choice. Psychological Research, 71, 42–63.
Hommel, B., & Colzato, L. (2004). Visual attention and the temporal dynamics of feature integration. Vis Cogn, 11, 483–521.
Hommel, B., & Colzato, L. (2009). When an object is more than a binding of its features: Evidence for two mechanisms of visual feature integration. Vis Cogn, 17, 120–140.
Hommel, B., Memelink, J., Zmigrod, S., & Colzato, L. S. (2013). Attentional control of the creation and retrieval of stimulus–response bindings. Psychological Research. doi:10.1007/s00426-012-0435-y
Hommel, B., Müsseler, J., Aschersleben, G., & Prinz, W. (2001). The Theory of Event Coding (TEC): A framework for perception and action planning. Behavioral and Brain Sciences, 24, 849–878, disc. 878–937. doi:10.1017/S0140525X01000103
Horner, A. J., & Henson, R. N. (2009). Bindings between stimuli and multiple response codes dominate long-lag repetition priming in speeded classification tasks. J Exp Psychol Learn Mem Cogn, 35, 757–779.
Horner, A. J., & Henson, R. N. (2011). Stimulus–response bindings code both abstract and specific representations of stimuli: Evidence from a classification priming design that reverses multiple levels of response representation. Mem Cogn, 39, 1457–1471. doi:10.3758/s13421-011-0118-8
Houghton, G., & Tipper, S. P. (1994). A model of inhibitory mechanisms in selective attention. In D. Dagenbach & T. H. Carr (Eds.), Inhibitory processes in attention, memory, and language (pp. 53–112). San Diego: Academic Press.
Ihrke, M., Behrendt, J., Schrobsdorff, H., Herrmann, J., & Hasselhorn, M. (2011). Response-retrieval and negative priming—Encoding- and retrieval-specific effects. Exp Psychol, 58, 154–161.
Jaeger, T. F. (2008). Categorical data analysis: Away from ANOVAs (transformation or not) and towards logit mixed models. J Mem Lang, 59, 434–446. doi:10.1016/j.jml.2007.11.007
Kahneman, D., & Treisman, A. (1984). Changing views of attention and automaticity. In R. Parasuraman & D. Davies (Eds.), Changing views of attention and automaticity (pp. 29–61). Orlando: Academic Press.
Kahneman, D., Treisman, A., & Gibbs, B. (1992). The reviewing of object files: Object-specific integration of information. Cogn Psychol, 24, 175–219. doi:10.1016/0010-0285(92)90007-O
Lassaline, M. E., & Logan, G. D. (1993). Memory-based automaticity in the discrimination of visual numerosity. J Exp Psychol Learn Mem Cogn, 19, 561–581. doi:10.1037/0278-7393.19.3.561
Ling, S., Liu, T., & Carrasco, M. (2009). How spatial and feature-based attention affect the gain and tuning of population responses. Vis Res, 49, 1194–1204.
Liu, T., Stevens, S., & Carrasco, M. (2007). Comparing the time course and efficacy of spatial and feature-based attention. Vis Res, 47, 108–113.
Logan, G. D. (1988). Toward an instance theory of automatization. Psychol Rev, 95, 492–527. doi:10.1037/0033-295X.95.4.492
Logan, G. D. (1990). Repetition priming and automaticity: Common underlying mechanisms? Cogn Psychol, 22, 1–35. doi:10.1016/0010-0285(90)90002-L
Logan, G. D. (1992). Shapes of reaction-time distributions and shapes of learning curves: A test of the instance theory of automaticity. J Exp Psychol Learn Mem Cogn, 18, 883–914. doi:10.1037/0278-7393.18.5.883
Logan, G. D. (1998). What is learned during automatization? II. Obligatory encoding of spatial location. J Exp Psychol Hum Percept Perform, 24, 1720–1736. doi:10.1037/0096-1523.24.6.1720
Logan, G. D. (2002). An instance theory of attention and memory. Psychol Rev, 109, 376–400.
Logan, G. D., & Etherton, J. L. (1994). What is learned during automatization? The role of attention in constructing an instance. J Exp Psychol Learn Mem Cogn, 20, 1022–1050. doi:10.1037/0278-7393.20.5.1022
Logan, G. D., Taylor, S. E., & Etherton, J. L. (1996). Attention in the acquisition and expression of automaticity. J Exp Psychol Learn Mem Cogn, 22, 620–638. doi:10.1037/0278-7393.22.3.620
Logan, G. D., Taylor, S. E., & Etherton, J. L. (1999). Attention and automaticity: Toward a theoretical integration. Psychological Research, 62, 165–181.
Logie, R. H., Brockmole, J. R., & Jaswal, S. (2011). Feature binding in visual short-term memory is unaffected by task-irrelevant changes of location, shape, and color. Mem Cogn, 39, 24–36. doi:10.3758/s13421-010-0001-z
Memelink, J., & Hommel, B. (2012). Intentional weighting: A basic principle in cognitive control. Psychological Research, 77, 249–259.
Moeller, B., & Frings, C. (2011). Remember the touch: Tactile distractors retrieve previous responses to targets. Exp Brain Res, 214, 121–130.
Moeller, B., Rothermund, K., & Frings, C. (2012). Integrating the irrelevant sound—Grouping modulates the integration of irrelevant auditory stimuli into event files. Exp Psychol, 59, 258–264.
Müller, H. J., Heller, D., & Ziegler, J. (1995). Visual search for singleton feature targets within and across feature dimensions. Percept Psychophys, 57, 1–17. doi:10.3758/BF03211845
Neill, W. T. (1997). Episodic retrieval in negative priming and repetition priming. J Exp Psychol Learn Mem Cogn, 23, 1291–1305. doi:10.1037/0278-7393.23.6.1291
Neill, W. T., Valdes, L. A., Terry, K. M., & Gorfein, D. S. (1992). Persistence of negative priming: II. Evidence for episodic trace retrieval. J Exp Psychol Learn Mem Cogn, 18, 993–1000. doi:10.1037/0278-7393.18.5.993
Neumann, E., & DeSchepper, B. (1992). An inhibition-based fan effect: Evidence for an active suppression mechanism in selective attention. Can J Psychol, 46, 1–40.
Posner, M. I., Snyder, C. R., & Davidson, B. J. (1980). Attention and the detection of signals. J Exp Psychol, 109, 160–174. doi:10.1037/0096-3445.109.2.160
R Development Core Team. (2013). R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. Retrieved from www.R-project.org/
Rothermund, K., Eder, A., & Frings, C. (2013). Integration of affective outcomes with actions and situations: Negative feedback prevents retrieval of response information from S–R episodes. Manuscript submitted for publication.
Rothermund, K., Wentura, D., & De Houwer, J. (2005). Retrieval of incidental stimulus–response associations as a source of negative priming. J Exp Psychol Learn Mem Cogn, 31, 482–495. doi:10.1037/0278-7393.31.3.482
Rugg, M. D., Fletcher, P., Frith, C., Frackowiak, R., & Dolan, R. (1997). Brain regions supporting intentional and incidental memory: A PET study. NeuroReport, 8, 1283–1287.
Tipper, S. (1992). Selection for action: The role of inhibitory mechanisms. Curr Dir Psychol Sci, 1, 105–109.
Treisman, A. (1992). Perceiving and re-perceiving objects. Am Psychol, 47, 862–875.
Treisman, A. (1996). The binding problem. Curr Opin Neurobiol, 6, 171–178. doi:10.1016/S0959-4388(96)80070-5
Treisman, A. M., & Gelade, G. (1980). A feature-integration theory of attention. Cogn Psychol, 12, 97–136. doi:10.1016/0010-0285(80)90005-5
Tukey, J. (1977). Exploratory data analysis. Reading: Addison-Wesley.
Tulving, E., Kapur, S., Craik, F., Moscovitch, M., & Houle, S. (1994). Hemispheric encoding/retrieval asymmetry in episodic memory: Positron emission tomography findings. Proc Natl Acad Sci, 91, 2016–2020.
van Dam, W., & Hommel, B. (2010). How object-specific are object files? Evidence for integration by location. J Exp Psychol Hum Percept Perform, 36, 1184–1192.
von der Malsburg, C. (1999). The what and why of binding: The modeler’s perspective. Neuron, 24, 95–104.
Waszak, F., & Pholulamdeth, V. (2009). Episodic S–R bindings and emotion: About the influence of positive and negative action effects on stimulus–response associations. Exp Brain Res, 194, 489–494.
Wiswede, D., Rothermund, K., & Frings, C. (2013). Not all errors are created equally: Specific ERN responses for errors originating from distractor-based response retrieval. Eur J Neurosci, 38, 3496–3506. doi:10.1111/ejn.12340
Wong, K. (2000). Dissociative prime–probe contextual similarity effects on negative priming and repetition priming: A challenge to episodic retrieval as a unified account of negative priming. J Exp Psychol Learn Mem Cogn, 26, 1411–1422.
Yantis, S., & Serences, J. (2003). Cortical mechanisms of space-based and object-based attentional control. Curr Opin Neurobiol, 13, 187–193.
Zmigrod, S., Spapé, M., & Hommel, B. (2009). Intermodal event files: Integrating features across vision, audition, taction, and action. Psychological Research, 73, 674–684.
Author note
The research reported in this article was supported by a grant from the Deutsche Forschungsgemeinschaft (No. FR 2133/1-2) to C.F. We thank Yaffa Yeshurun, Iring Koch, Gordon Logan, and an anonymous reviewer for their helpful feedback on previous versions of this article.
Author information
Authors and Affiliations
Corresponding author
Appendix
Appendix
With respect to the problems of using categorical outcome variables in ANOVA (see, e.g., Dixon, 2008; Jaeger, 2008), all error analyses were also analyzed using multilevel logistic regression, with trials at Level 1 and participants as Level 2 variables (Baayen, Davidson, & Bates, 2008). The factors Response Repetition, Distractor Repetition, and Attention Modulation, as well as all possible interaction effects, were modeled as random factors (i.e., they were allowed to vary across participants). Analyses were computed using the R software (R Development Core Team, 2013) and the R package lme4 (Bates, Maechler, Bolker, & Walker, 2013). Table A1 reports the intercepts, z values, and p values of all main and interaction effects of all experiments. Taken together, one can say that the data pattern mimics the data pattern using MANOVA in the main text for Experiments 1a and 3; for Experiments 1b and 2, the model failed to converge after 10,000 evaluations.
Rights and permissions
About this article
Cite this article
Moeller, B., Frings, C. Attention meets binding: Only attended distractors are used for the retrieval of event files. Atten Percept Psychophys 76, 959–978 (2014). https://doi.org/10.3758/s13414-014-0648-9
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13414-014-0648-9