A Review of the Immunomodulating Components of Maternal Breast Milk and Protection Against Necrotizing Enterocolitis


1
Department of Pediatrics, Division of Newborn Medicine, Washington University School of Medicine, St. Louis, MO 63110, USA
2
University of Pittsburgh School of Medicine, Medical Scientist Training Program, Pittsburgh, PA 15213, USA
*
Author to whom correspondence should be addressed.
Nutrients 2020, 12(1), 14; https://doi.org/10.3390/nu12010014
Submission received: 12 November 2019
/
Revised: 5 December 2019
/
Accepted: 17 December 2019
/
Published: 19 December 2019
(This article belongs to the Special Issue Neonatal Nutrition for Inflammatory Disorders and Necrotizing Enterocolitis)
Abstract
:Breast milk contains immunomodulating components that are beneficial to newborns during maturation of their immune system. Human breast milk composition is influenced by an infant’s gestational and chronological age, lactation stage, and the mother and infant’s health status. Major immunologic components in human milk, such as secretory immunoglobulin A (IgA) and growth factors, have a known role in regulating gut barrier integrity and microbial colonization, which therefore protect against the development of a life-threatening gastrointestinal illness affecting newborn infants called necrotizing enterocolitis (NEC). Breast milk is a known protective factor in the prevention of NEC when compared with feeding with commercial formula. Breast milk supplements infants with human milk oligosaccharides, leukocytes, cytokines, nitric oxide, and growth factors that attenuate inflammatory responses and provide immunological defenses to reduce the incidence of NEC. This article aims to review the variety of immunomodulating components in breast milk that protect the infant from the development of NEC.
1. Introduction
Necrotizing enterocolitis (NEC) is a severe gastrointestinal disease in preterm infants with associated mortality as high as 50% in cases that require surgical intervention [1]. NEC occurs in 1–5% of patients admitted to a neonatal intensive care unit (NICU), and increased incidence and fatality occurs in infants with prematurity and low birth weight [2]. The etiology of NEC is complex, and pathogenesis is attributed to inflammation of the neonatal gastrointestinal tract by triggers such as commercial formula feeds, intestinal dysbiosis, and immaturity of gut mucosal immunity. Treatment of NEC requires withholding enteral feeds and potent antimicrobial agents, and these infants are at risk of adverse long term outcomes. Identification of factors that contribute to the prevention of NEC remains a high priority in neonatal research. There is a consensus regarding the protective nature of breast milk in the prevention of NEC development. Human breast milk, in contrast to commercial formulas, contains soluble and cellular components that provide infants with passive immunity to their gastrointestinal tract. These antimicrobial and bioactive factors are multi-functional and anti-inflammatory, with an established protective role against the development of NEC. An early prospective study of 926 infants showed that exclusively formula-fed preterm infants were six to ten times more likely to acquire NEC as compared to preterm infants nourished with human milk alone [3]. Furthermore, an analysis of 243 infants in a randomized trial showed that preterm infants less than 30 weeks’ gestation who received maternal milk had reduced incidence of late-onset sepsis or NEC as compared with preterm infants who received donor breast milk or commercial formula [4]. Sullivan and colleagues demonstrated in a randomized, controlled, multicenter trial that extremely premature infants who received only human milk, including human milk-derived fortification, had decreased rates of NEC compared to those infants exposed to bovine milk-derived products with a number needed to treat of 10 infants to prevent one NEC case [5].
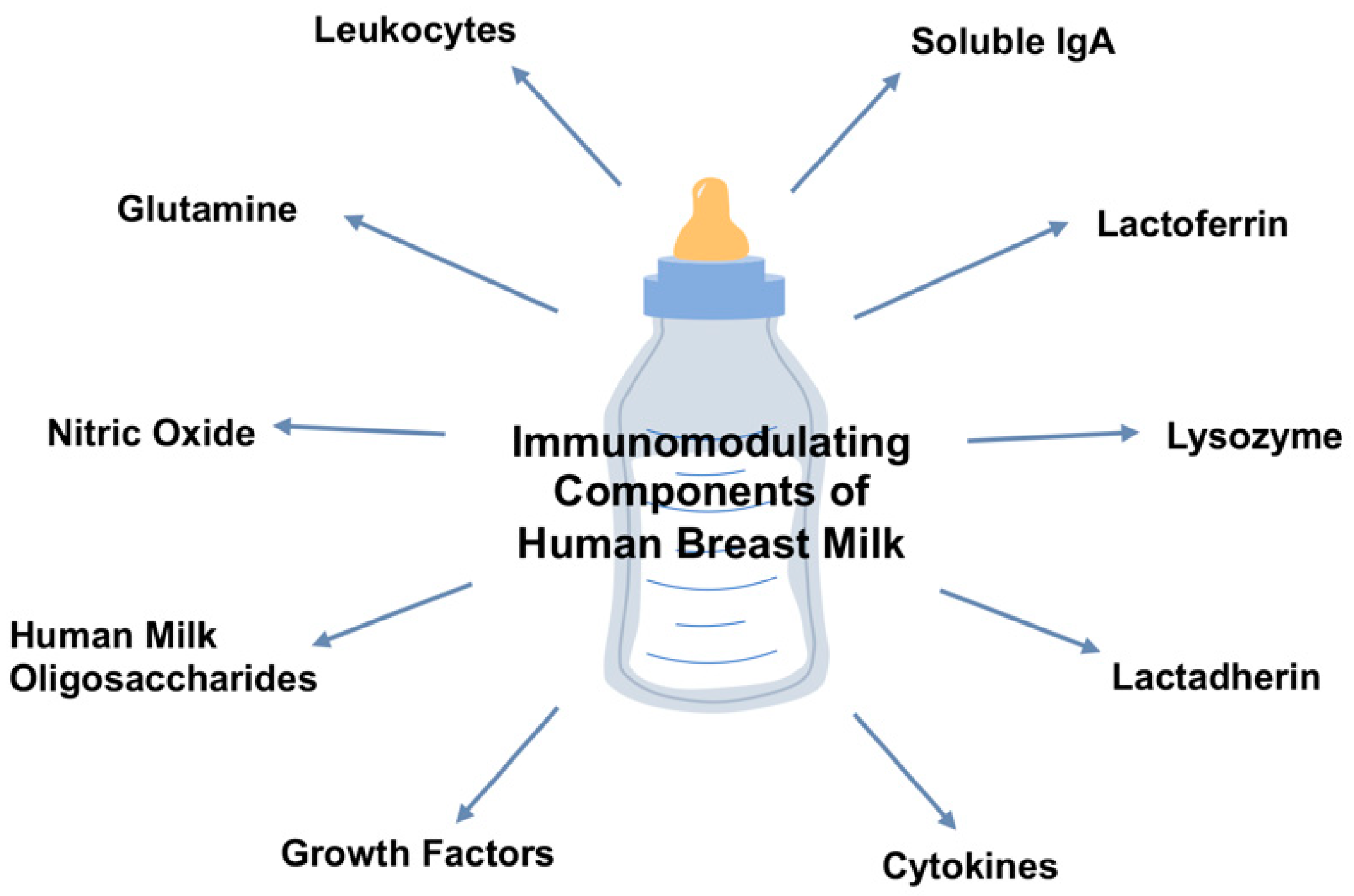
Breast milk composition is complex, dynamic, and influenced by a variety of maternal factors. Immunoglobulins, antimicrobial peptides, growth factors, human milk oligosaccharides, cytokines, L-glutamine, and nitric oxide in breast milk maintain roles in the enhancement of the neonatal intestinal barrier function and the reduction of NEC. This article aims to review the protective role of breast milk and its components against NEC, as shown in Figure 1.
2. Breast Milk and the Host-Microbial Relationship
2.1. Maternal Soluble IgA
Bioactive components of maternal milk, particularly immunoglobulin A (IgA), have known influential effects on the neonatal microbiota. IgA is the most plentiful antibody in human milk and comprises a significant portion of total protein content in colostrum [6]. IgA produced by the maternal mammary glands undergoes proteolytic cleavage to release secretory IgA (sIgA), permitting transport into human milk [7]. Breast milk sIgA provides critical antimicrobial defense to the neonatal gastrointestinal tract through inhibition of pathogen attachment to mucosal surfaces, neutralization of microbial toxins, and provision of passive immunity. IgM and IgG are of lesser abundance in human milk but also have known immune-surveillance properties.
The sIgA levels in breast milk decrease over time during the postpartum period [8]. Recent studies have identified no difference in breast milk sIgA concentration of preterm versus term breast milk [6,8], although Mehta and Petrova identified that preterm milk contains a higher concentration of sIgA in the first six to eight days of lactation [9]. Breast milk sIgA is a critical component, as it promotes colonization of commensal microbiota, decreasing the activity of pattern recognition receptors and subsequent downstream inflammation within the intestinal epithelium [10]. A humoral response with increased levels of sIgA in mature breast milk has been shown to occur in response to an infection in the mother or infant [11]. Gopalakrishna and colleagues studied the role that sIgA in breast milk plays in the pathogenesis of NEC [12]. They determined the proportion of IgA-associated intestinal bacteria and discovered that premature infants with an exclusive formula-fed diet contained very low levels of IgA-associated intestinal bacteria. Furthermore, infants with NEC had higher levels of IgA-unbound Enterobacteriaceae when compared with healthy age-matched controls. This suggests that insufficient concentrations of IgA and decreased IgA-bound bacteria in the intestine may be causative factors of insufficient microbiome diversity and increased risk of NEC development [12]. In a neonatal mouse model of NEC, pups reared by IgA-deficient mothers showed susceptibility to intestinal disease despite receiving maternal milk, suggesting that maternal IgA can define the host-microbiota relationship and underscoring that the IgA in milk plays an important role in the susceptibility to NEC [12].
2.2. Lactoferrin
Lactoferrin is an abundant peptide in human milk and has known roles in host defense and antimicrobial properties. When lactoferrin encounters proteolysis in acidic conditions, such as in the stomach, lactoferricin is produced. Lactoferricin has both strong antibacterial and some antiviral activity with immunomodulatory capabilities [13]. In particular, human lactoferricin has a potent ability to neutralize endotoxin activity, prevent activation of mononuclear cells, and ultimately prevent the secretion of cytokines, such as interleukin (IL)-1β, IL-6, tumor necrosis factor (TNF)-α, and IL-8, that contribute to inflammation [13,14]. Togawa and colleagues demonstrated that the administration of enteral lactoferrin in rats attenuated colonic inflammation after induction of colitis [15]. Many studies have subsequently evaluated the modulatory role of lactoferrin in antimicrobial and immunological defenses in infants. A Cochrane Review of six, small, randomized control trials (RCT) that provided lactoferrin supplementation to enteral feeds found decreased late-onset bacterial and fungal sepsis in preterm infants, although the evidence was identified as low in quality [16]. A systematic review and meta-analysis reviewed nine RCTs and showed that prophylactic lactoferrin significantly reduced the incidence of late-onset sepsis and NEC (Bell’s stage II or greater) [17]. Most recently, a large randomized control trial of 2203 infants contrasted these prior findings, demonstrating that enteral supplementation with bovine lactoferrin did not reduce NEC or the incidence of infection or mortality [18].
2.3. Lysozyme
Lysozyme, as an immune-active enzyme in colostrum and breast milk, has many bactericidal effects. In synergy with lactoferrin, lysozyme can bind to lipopolysaccharide (LPS) on outer bacterial membranes, which provides lysozyme access to degrade internal proteoglycan matrices of bacterial membranes. Studies of breast milk composition have shown that premature breast milk as compared to term breast milk has higher lysozyme content [6,9], although other studies have found no difference [8]. In the gastrointestinal tract, Paneth cells within the crypts of Lieberkühn produce a variety of antimicrobial peptides, including lysozyme, which are secreted in response to enteric pathogens [19]. Of relevance to NEC, in the small intestinal biopsies of premature infants with NEC, there were decreased concentrations of Paneth cells compared to controls [20]. The role of lysozyme has been studied in a mouse model of NEC utilizing Paneth cell ablation. This model consists of 14-day-old pups treated with dithizone, a heavy metal chelator, followed by luminal infection with Klebsiella [21]. The pups in this experimental group developed a NEC-like injury, suggesting the significance of lysozyme and antimicrobial protection provided by Paneth cells can regulate the inflammatory response in NEC [21]. A subsequent study using an experimental murine NEC model demonstrated that Paneth cell deficiency induces a disruption in the intestinal microbiome, and in particular, the development of an Enterobacteriaceae bloom, which has been shown to precede NEC in humans [22]. These results signify the potential significance of breast milk lysozyme in protecting breast fed infants from the intestinal inflammatory insult seen in NEC.
2.4. Lactadherin
Lactadherin (milk fat globule-epidermal growth factor (EGF) factor VIII) is a human milk glycoprotein that contributes to apoptotic cell phagocytosis [23]. A deficiency of lactadherin has been strongly associated with inflammatory and autoimmune diseases and has been shown to maintain homeostasis of the intestinal epithelium through the migration of epithelial cells. In a model of seven-week-old mice, treatment with recombinant lactadherin resulted in protection from colitis, as demonstrated by downregulation of pro-inflammatory cytokines and improved histological scores [23]. Additionally, in a neonatal rat model of NEC-like intestinal injury, supplementation with recombinant human lactadherin attenuated the disruption of cellular tight junctions [24].
2.5. Epidermal Growth Factor
The growth factors in breast milk serve a protective role in helping to facilitate the intestinal mucosal barrier maturation. Maternal milk and colostrum contain epidermal growth factor (EGF) and are the predominant sources of intestinal EGF during the postnatal phase. The roles of EGF in the development of the intestine, as well as the response and repair of the intestine during intestinal injury or infection, have been reported [25]. EGF levels are decreased in the saliva and serum of premature infants with NEC when compared to infants without NEC. In a study of salivary EGF, infants with NEC had lower salivary EGF in the first week after birth and greater increases from week of life one to two as compared to infants without NEC, suggesting that NEC development may be attributed to overall lower EGF concentrations in the at-risk neonate [26]. EGF also has proposed effects on goblet cells and the production of mucin in the intestinal epithelium. Clark and colleagues showed that treatment with EGF resulted in an increased number of goblet cells and increased the production of mucin in the small intestine [27].
NEC has been associated with impaired intestinal barrier function and epithelial cell apoptosis. The in vivo treatment with enteral EGF has shown to regulate the expression of tight junction proteins, occludin and claudin-3 as well as normalize their expression at the site of NEC injury, helping to maintain the gut barrier [27]. Additionally, enteral EGF administration can increase expression of the anti-apoptotic protein, Bcl-2, and decrease levels of the pro-apoptotic protein, Bax. The role of EGF in balancing apoptosis regulators provides implications of an opportunity for future therapeutic strategies to protect the intestinal barrier from injury in NEC [28,29].
2.6. Heparin-Binding Epidermal Growth Factor
The developing fetus and the breast fed newborn are continually exposed to Heparin-binding epidermal growth factor (HB-EGF), which is present in both amniotic fluid and breast milk, suggesting its possible role in gastrointestinal epithelium development both in utero and during the neonatal period [30]. As a member of the EGF family, HB-EGF binds to the EGF receptor (EGFR) and has known mitogenic effects. HB-EGF is expressed in response to hypoxia, tissue damage, and oxidative stress, including in the intestine, and has a pivotal role in tissue regeneration and repair [31,32]. In seeking to evaluate the role of exogenous HB-EGF in the context of NEC, Dvorak and colleagues demonstrated that either the oral administration of HB-EGF or EGF significantly reduced NEC in a premature rat model through increased production of MUC2, a secretory mucin [33]. However, the concurrent administration of both growth factors did not confer better protection and physiologic doses of EGF provided better protection [33]. In another study, enteral administration of HB-EGF to neonatal rat pups decreased the incidence and severity of NEC and reduced intestinal permeability as demonstrated by a low serum concentration of enterally-administered fluorescein isothiocyanate-dextran [32]. The results of these studies suggest a potential role of HB-EGF in the attenuation of intestinal injury during NEC.
2.7. Transforming Growth Factor-β2
Human milk contains high concentrations of the transforming growth factor-β isoform, transforming growth factor-β2 (TGF-β2), which has immunomodulatory effects on intestinal maturation, immunoglobulin production, and a suppressive effect on T cells [34]. Breast milk with higher concentrations of TGF-β2 is associated with a higher diversity of intestinal microbial composition in the neonate, a factor that is known to lower the risk of adult immunological diseases [34]. Of note, preterm human milk has been shown to have reduced TGF-β bioactivity [35]. Maheshwari and colleagues analyzed TGF-β2 expression in premature infant intestinal tissue samples and observed lower TGF-β2 expression and bioactivity in patients with NEC as compared with controls [36]. In a murine experimental model of NEC, enterally administered recombinant TGF-β2 showed protective effects against NEC-like mucosal injury [36]. The addition of recombinant TGF-β2 to milk has been investigated as a preventative strategy to boost the anti-inflammatory properties of milk and prevent the development of NEC. However, it was discovered that in human preterm milk, TGF-β2 is sequestered by chondroitin sulfate proteoglycans, which therefore inhibits its biological activity [35]. Consequently, the digestion of human preterm milk with chondroitinase resulted in the activation of endogenous TGF-β2 and also restored the bioactivity of recombinant TGF-β2 [35]. These findings suggest chondroitinase digestion of preterm milk may be an option for preventing NEC by enhancing the anti-inflammatory properties of the milk.
2.8. Prebiotics and Oligosaccharides
Human milk oligosaccharides (HMOs) are complex sugars present in high abundance in breast milk. HMOs serve as prebiotics and metabolic substrates with targeted antimicrobial activity, allowing beneficial bacteria to thrive while suppressing those which are potentially harmful [37,38]. In an in vitro epithelial model of the crypt-villus axis, treatment with HMOs resulted in reduced intestinal cell proliferation, but promoted epithelial cell differentiation, indicating a potential role in intestinal maturation [39].
HMOs ingested from breast milk undergo only minimal degradation in the infant’s acidic stomach and by the pancreatic and brush border enzymes in order to reach the distal small intestine and colon [37,38]. Preclinical animal studies as well as human studies in mother–infant dyads support the contributions of HMOs in reduction of the development of NEC. In a cohort study comprised of 200 mother–infant dyads, the composition of HMOs in breast milk was analyzed [40]. One specific HMO, disialyllacto-N-tetraose (DSLNT) was identified to be present in significantly lower concentrations in those infants who developed NEC [40]. Measurement of DSLNT levels in maternal milk may therefore provide additional insight into why some breast fed infants are still at risk of NEC.
Enteral administration with supplemental HMOs have been studied as potential therapeutics in reducing the risk of NEC [41,42]. For example, in a neonatal rat model of NEC, animals were fed with DSLNT-containing formula, which resulted in reduced severity of NEC based on pathology scores and improved survival [43]. The same study showed that galacto-oligosaccharides, an infant formula additive, similar although structurally different from HMOs, demonstrated no effect on NEC severity or survival in neonatal rats [43]. In a preterm pig model of NEC, receiving supplemental feeds with a mixture of HMOs have not shown a significant difference in NEC severity, gut microbial colonization, or intestinal permeability [42]. However, in a neonatal mouse model of NEC, enteral administration of another HMO found in breast milk, 2’fucosyllactose (HMO-2’FL), resulted in the preservation of mesenteric perfusion and restored the expression of endothelial nitric oxide synthase (eNOS), a vasodilatory molecule necessary for intestinal perfusion [44]. The results of these studies suggest key roles of HMO-2’FL and DSLNT as protective components of breast milk in the prevention of NEC development.
2.9. Glutamine
Free amino acids comprise 3–5% of the total amino acids in human milk [45]. In a longitudinal analysis of breast milk from healthy mothers of term infants, glutamine and glutamic acid were among the most plentiful free amino acids in the first three months of lactation [45]. Levels of glutamine increased significantly during the first to the third month of lactation [45,46]. In addition, breast milk, which contains higher concentrations of glutamine, amongst other free amino acids, is associated with more rapid weight gain [46] and increased length [47] in the infant. Glutamine also holds a relevant role in maintaining gut barrier integrity. For example, glutamine augments the effects of growth factors and influences cell signaling pathways involved in intestinal cell proliferation and differentiation, as well as the expression of tight junctions [48,49]. Glutamine has also exhibited anti-apoptotic properties in intestinal cells, attributed to its role in the production of glutathione [48,49].
Neonates deficient in circulating amino acids such as glutamine and arginine, are associated with a higher risk of NEC development [50]. In a neonatal rat model of NEC, pups receiving exogenous administration of glutamine had reduced pathology injury scores and reduced ileal mRNA expression of the innate immune receptors, Toll-like receptor (TLR)-2 and TLR-4 [51]. As TLR-2 and TLR-4 have established roles in inducing synthesis of inflammatory mediators and increasing apoptosis in NEC [52,53,54], their reduced expression by glutamine supplementation suggests a mechanism by which it mediates protection. In a small study that evaluated the outcomes of arginine and glutamine supplementation in 25 preterm neonates of less than 34 weeks’ gestation, there were no infants that developed NEC in the glutamine group and no difference in the NEC incidence in the arginine group [55]. However, large RCTs of infants diagnosed with severe gastrointestinal disease, including NEC, spontaneous intestinal perforation, and intestinal structural anomalies, did not show a decreased risk of death or severe infections while receiving enteral glutamine [56]. Additional large RCTs evaluating glutamine supplementation in preterm infants did not show a benefit in decreasing the risk of death, intestinal disease, or long term developmental outcomes [57]. Therefore, despite the significant levels of glutamine in human breast milk, there is insufficient evidence for exogenous supplementation of glutamine as a preventative strategy for NEC at this time.
3. Breast Milk and Immune Homeostasis
3.1. Cellular Mechanisms
There are two primary pathways for maternal cellular transfer to the infant—placental transmission and oral transmission through breastfeeding [58]. Breast milk leukocytes, including macrophages and neutrophils, survive passage through the neonatal gastrointestinal tract and translocate to blood, lymph nodes, spleen, and liver [11,58,59]. Understanding the physiological significance of the transfer of human milk cells to neonates can provide insight into the protective properties of breast milk on the infant recipient.
The progression through maturational stages of lactation involves alterations in breast milk leukocyte composition and concentration. In an analysis of CD45+ leukocytes in breast milk, colostrum contained the highest number of leukocytes compared with transitional milk (8–12 days postpartum) and mature milk (26–30 days postpartum) [60]. The infant’s gestational age at birth is also associated with changing concentrations of certain types of breast milk leukocytes. Colostrum contains lower levels of non-cytotoxic T cells and B lymphocytes with increased gestational age whereas mature milk of preterm mothers contains lower cytotoxic T cell and natural killer (NK) cell levels when compared to term milk [60]. In seeking to understand the impact of maternal milk leukocytes on the breastfeeding infant, Cabinian and colleagues used a murine model to examine the transport and survival of maternal breast milk leukocytes, primarily T cells, to the gastrointestinal Peyer’s patches of the suckling pup [61]. The observed transfer of cells to the Peyer’s patches implicates the role of breast milk leukocytes in neonatal intestinal development and localized immunological maturation. The overall relevance of the differences in human milk cellular content and transfer on the development of NEC requires further study.
Additionally, maternal and infant bacterial infections influenced concentrations of breast milk leukocytes and cytokines, notably macrophages and TNF-α levels [11,62]. Maternal infection can induce a significant leukocyte surge that ranges from 0.7% to 93.6% of total cells in breast milk [11]. A smaller increase in breast milk leukocytes has been observed when the breastfeeding infant develops an infection [11]. Riskin and colleagues identified that macrophages, as well as neutrophils, comprise the majority of breast milk leukocytes in mothers with a sick infant [62]. The increase in breast milk leukocytes in response to an inflammatory process in the mother/infant dyad suggests a dynamic interaction between maternal and infant immune systems and further supports the benefits of breast milk.
3.2. Cytokines
Preterm infants, when compared with their term counterparts, exhibit immune immaturity, which includes lower production of cytokines and other immunological proteins during challenge with an inflammatory insult [59]. The presence of cytokines in breast milk provides passive protection and immune modulation in the infant recipient and results in absorption into the systemic circulation. In particular, these cytokines include IL-1, IL-2, IL-6, IL-8, IL-10, interferon (IFN)-γ, and TNF-α (Table 1). Breast milk produced by mothers of full-term infants contains high levels of IL-2, IL-8, and IL-10, with levels decreasing drastically by day 21 of lactation. In contrast, mothers of preterm infants have significantly lower levels of cytokines in the colostrum when compared to mothers of full-term infants [63].
High levels IL-1RA, an IL-1 receptor, have been detected in breast milk [64]. IL-1β is a member of the IL-1 family and is known to induce an endogenous innate inflammatory response in enterocytes, upregulate expression of pro-inflammatory IL-8, and stimulate the nuclear factor kappa beta (NF-kB) pathway. However, human milk has demonstrated the ability to attenuate the IL-1β-dependent activation of IL-8 [64]. The protective effects of breast milk on suppressing this NF-kB-mediated pro-inflammatory immune response has been shown in intestinal epithelial cells both in vitro and in vivo, providing evidence of a mechanism mediating protection against NEC development [53,64].
Neonates have a known deficiency in the production of IL-2, which is a necessary cytokine in the recruitment of T cells required to produce an antigen-specific immune response [65,66]. Human milk, therefore, provides an ideal source of IL-2 for the newborn. Levels of aqueous IL-2 in human milk are of highest concentration in colostrum with reduced levels in later stages of lactation [63,65]. The presence of IL-2 in breast milk, which is absorbed by the gastrointestinal tract of the infant, may enter the systemic circulation to influence the maturing immune system.
IL-6 is a pro-inflammatory cytokine in the acute phase of the inflammatory response [59]. Multiple studies have observed high levels of IL-6 in colostrum [63,67,68]. An early analysis of IL-6 in breast milk showed that the presence of an anti-IL-6 antibody in colostrum caused decreased production of IgA by mononuclear leukocytes, suggesting a relationship between IL-6 and IgA production in breast milk [67]. Ustundag and colleagues noted higher levels of breast milk IL-6 at two weeks postpartum in mothers of term infants when compared to milk from mothers of preterm infants [63]. The prevalence of IL-6 in breast milk with uptake by the infant recipient may have a significant biologic role in neonatal immune homeostasis.
IL-8 expression by macrophages, endothelial cells, and epithelial cells provides chemotactic activity for a neutrophil-dependent response to acute inflammation, such as in sepsis and NEC. A decline in IL-8 levels in breast milk occurs with the advancement in lactational stage [69,70]. Although one study found no difference in breast milk IL-8 expression in mothers of infants of different gestational ages [63], others have identified higher IL-8 levels in the breast milk of mothers of preterm infants [69]. Maheshwari and colleagues showed that fetal and adult human intestinal cells treated with recombinant IL-8 in vitro had increased cell proliferation and differentiation [69]. Additionally, intestinal cells exposed to injury in vitro demonstrated increased viability when treated with recombinant IL-8 [69]. Thus, the dynamic effects of IL-8 on the developing intestine suggests its physiologic role in intestinal development as a component of human breast milk.
The anti-inflammatory properties of IL-10 attenuate the immune response to an infection and maintain tissue homeostasis by inhibition of the activity of Th1 effector cells, NK cells, and macrophages [71,72]. IL-10 can inhibit the production of inflammatory IL-1, IL-6, IL-8, and TNF-α [72,73]. One study found IL-10 in the aqueous and non-aqueous phases of human milk, with concentrations found to be highest within the first 24 hours of lactation [72]. IL-10 in breast milk affects the infant by attracting CD8+ T lymphocytes [72,74] and promoting immunoglobulin synthesis by B cells [72,75].
IFN-γ is a pro-inflammatory cytokine found in human milk in low concentrations with decreasing levels in the months following birth [76]. IFN-γ is secreted by activated T cells and NK cells and enhances intestinal macrophage activation [76]. IFN-γ is involved in the signaling pathways that increase intestinal epithelial barrier permeability [77] and is also detected in higher concentrations in the ileum of patients with NEC [78]. As infants have a reduced ability to produce IFN-γ due to an immature immune system [79], breast milk may provide the infant with IFN-γ and other pro-inflammatory cytokines needed to produce a host defense response against inflammation or infection.
Neonates are deficient in the production of the pro-inflammatory cytokine TNF-α and its receptors, TNF-α receptor I and II, increasing susceptibility to infection due to immune cell dysregulation. TNF-α is produced by a variety of immune cell types, including granulocytes and CD4+ lymphocytes. TNF-α, as an endogenous pyrogen, contributes to systemic inflammation and immune cell regulation. One study quantified the amount of detectable TNF-α in breast milk and colostrum and identified the majority of TNF-α to be in association with its soluble receptor [80]. The low amount of unbound TNF-α in breast milk was theorized to decrease TNF-α pro-inflammatory bioactivity [80]. Two additional studies have identified a significantly decreased amount of TNF-α in colostrum of mothers who delivered very preterm (less than 30 weeks’ gestation) when compared to term and preterm groups, suggesting one reason why preterm infants have increased susceptibility to infection and impaired immunity [63,81].
Overall, the evidence suggests that infants, particularly those born preterm, have insufficient ability to mount an adequate immunological defense due to reduced production of a variety of cytokines. Breast milk can therefore supplement infants with maternal cytokines that may provide immune benefits in the protection against neonatal inflammatory diseases such as NEC.
3.3. Nitric Oxide
Nitric oxide (NO) is a soluble molecule produced by isoforms of NO synthase (NOS) and serves as a potent vasodilator and neurotransmitter at low, physiologic levels [82,83]. Infants derive NO from dietary sodium nitrite, which is then converted to NO within the gastrointestinal tract by commensal microbes [82,83]. NO is present in breast milk, as shown in an analysis of healthy lactating mothers evaluating the concentration of breast milk nitric oxide concentration on postpartum days one through five [84]. Exclusively breastfeeding mothers had significantly higher nitric oxide concentrations in their milk, compared with milk expressed from mothers who decided to exclusively formula-feed their infant [84]. It was theorized that infant suckling activates NOS within the mammary gland with subsequent secretion of NO into breast milk, which then confers protection to the intestine of the infant through regulation of intestinal blood flow and maintenance of vascular tone [84].
The upregulation of inducible NOS (iNOS) in response to the release of cytokines and growth factors has been implicated in NEC pathogenesis [82,85]. During inflammation, high levels of NO, and its derivative peroxynitrite, contribute to epithelial damage and the disruption of the integrity of the intestinal barrier [82]. To study abnormal NOS signaling in NEC, Yazji and colleagues used a murine model of NEC and selectively deleted endothelial TLR-4 expression, which subsequently resulted in impaired microvascular intestinal perfusion, increased severity of NEC, and reduced endothelial NOS (eNOS) expression [86]. Additionally, as compared with commercial formula, breast milk was identified to have higher levels of sodium nitrate, which serves as a precursor for nitrite and nitric oxide. Enteral administration of exogenous sodium nitrate was associated with decreased severity of NEC and improved intestinal perfusion [86]. Overall, these results suggest the protective role of breast milk in augmenting physiologic nitrate-nitrite-NO signaling to improve intestinal vascular perfusion and protect against intestinal barrier disruption in NEC.
4. Summary
Human breast milk contains a dynamic diversity of bioactive components needed for infant growth, immune homeostasis, and intestinal maturation. The composition of human milk varies with the stage of lactation, gestational age of the infant, the health of the mother/infant dyad, and the nutritional status of the mother. The dietary intake of the breastfeeding mother has been shown to influence the variability of human milk concentrations of fat-soluble and water-soluble vitamins and other nutrients. These nutrients, including immunoglobulins, growth factors, cytokines, and immune cells, have been demonstrated to transfer from the mother to the neonate through breast milk [87,88]. The ability of these components to regulate intestinal cell proliferation and differentiation as well as influence gut microbial colonization emphasizes the protective role of breast milk in infant metabolism and neurodevelopment, intestinal microbial homeostasis, and protection against NEC [87,88]. The growing field of research studying the outcomes related to breastfeeding reinforces the immunological value of breast milk on infant nutrition and protection from inflammatory disorders such as NEC.
Author Contributions
L.S.N., O.B.P. and M.G. contributed to the writing—Original draft preparation, review and editing of the initial version manuscript. L.S.N. and M.G. edited and revised the manuscript. L.S.N., O.B.P. and M.G. approved the final version of the manuscript. All authors have read and agreed to the published version of the manuscript.
Acknowledgments
MG is supported by R01DK118568 from the National Institutes of Health, March of Dimes Foundation Grant No. 5-FY17-79, the St. Louis Children’s Hospital Foundation, the Children’s Discovery Institute of Washington University and St. Louis Children’s Hospital, and the Department of Pediatrics at Washington University School of Medicine, St. Louis. MG has previously received sponsored research agreement funding from Astarte Medical Partners, and participated in a neonatal microbiome advisory board for Abbott Laboratories. None of the above sources had any role in this study.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the writing of this manuscript.
References
- Neu, J.; Walker, W.A. Necrotizing enterocolitis. N. Engl. J. Med. 2011, 364, 255–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, P.W.; Stoll, B.J. Necrotising enterocolitis. Lancet 2006, 368, 1271–1283. [Google Scholar] [CrossRef]
- Lucas, A.; Cole, T.J. Breast milk and neonatal necrotising enterocolitis. Lancet 1990, 336, 1519–1523. [Google Scholar] [CrossRef]
- Schanler, R.J. Randomized trial of donor human milk versus preterm formula as substitutes for mothers’ own milk in the feeding of extremely premature infants. Pediatrics 2005, 116, 400–406. [Google Scholar] [CrossRef]
- Sullivan, S.; Schanler, R.J.; Kim, J.H.; Patel, A.L.; Trawöger, R.; Kiechl-Kohlendorfer, U.; Chan, G.M.; Blanco, C.L.; Abrams, S.; Cotten, C.M.; et al. An exclusively human milk-based diet is associated with a lower rate of necrotizing enterocolitis than a diet of human milk and bovine milk-based products. J. Pediatr. 2010, 156, 562–567.e1. [Google Scholar] [CrossRef] [Green Version]
- Trend, S.; Strunk, T.; Lloyd, M.L.; Kok, C.H.; Metcalfe, J.; Geddes, D.T.; Lai, C.T.; Richmond, P.; Doherty, D.A.; Simmer, K.; et al. Levels of innate immune factors in preterm and term mothers’ breast milk during the 1st month postpartum. Br. J. Nutr. 2016, 115, 1178–1193. [Google Scholar] [CrossRef] [Green Version]
- Rogier, E.W.; Frantz, A.L.; Bruno, M.E.C.; Wedlund, L.; Cohen, D.A.; Stromberg, A.J.; Kaetzel, C.S. Secretory antibodies in breast milk promote long-term intestinal homeostasis by regulating the gut microbiota and host gene expression. Proc. Natl. Acad. Sci. USA 2014, 111, 3074–3079. [Google Scholar] [CrossRef] [Green Version]
- Hsu, Y.C.; Chen, C.H.; Lin, M.C.; Tsai, C.R.; Liang, J.T.; Wang, T.M. Changes in preterm breast milk nutrient content in the first month. Pediatr. Neonatol. 2014, 55, 449–454. [Google Scholar] [CrossRef] [Green Version]
- Mehta, R.; Petrova, A. Biologically active breast milk proteins in association with very preterm delivery and stage of lactation. J. Perinatol. 2011, 31, 58–62. [Google Scholar] [CrossRef] [Green Version]
- Rogier, E.W.; Frantz, A.L.; Bruno, M.E.C.; Kaetzel, C.S. Secretory IgA is concentrated in the outer layer of colonic mucus along with gut bacteria. Pathogens 2014, 3, 390–403. [Google Scholar] [CrossRef] [Green Version]
- Hassiotou, F.; Hepworth, A.R.; Metzger, P.; Tat Lai, C.; Trengove, N.; Hartmann, P.E.; Filgueira, L. Maternal and infant infections stimulate a rapid leukocyte response in breastmilk. Clin. Transl. Immunol. 2013, 2, e3. [Google Scholar] [CrossRef] [PubMed]
- Gopalakrishna, K.P.; Macadangdang, B.R.; Rogers, M.B.; Tometich, J.T.; Firek, B.A.; Baker, R.; Ji, J.; Burr, A.H.P.; Ma, C.; Good, M.; et al. Maternal IgA protects against the development of necrotizing enterocolitis in preterm infants. Nat. Med. 2019, 25, 1110–1115. [Google Scholar] [CrossRef]
- Gifford, J.L.; Hunter, H.N.; Vogel, H.J. Lactoferricin: A lactoferrin-derived peptide with antimicrobial, antiviral, antitumor and immunological properties. Cell. Mol. Life Sci. 2005, 62, 2588–2598. [Google Scholar] [CrossRef] [PubMed]
- Palmeira, P.; Carneiro-Sampaio, M. Immunology of breast milk. Rev. Assoc. Med. Bras. 2016, 62, 584–593. [Google Scholar] [CrossRef]
- Togawa, J.I.; Nagase, H.; Tanaka, K.; Inamori, M.; Nakajima, A.; Ueno, N.; Saito, T.; Sekihara, H. Oral administration of lactoferrin reduces colitis in rats via modulation of the immune system and correction of cytokine imbalance. J. Gastroenterol. Hepatol. 2002, 17, 1291–1298. [Google Scholar] [CrossRef]
- Pammi, M.; Suresh, G. Enteral lactoferrin supplementation for prevention of sepsis and necrotizing enterocolitis in preterm infants. Cochrane Database Syst. Rev. 2017, 2017, CD007137. [Google Scholar] [CrossRef]
- He, Y.; Cao, L.; Yu, J. Prophylactic lactoferrin for preventing late-onset sepsis and necrotizing enterocolitis in preterm infants. Medicine (Baltimore) 2018, 97, e11976. [Google Scholar] [CrossRef]
- Griffiths, J.; Jenkins, P.; Vargova, M.; Bowler, U.; Juszczak, E.; King, A.; Linsell, L.; Murray, D.; Partlett, C.; Patel, M.; et al. Enteral lactoferrin to prevent infection for very preterm infants: The ELFIN RCT. Health Technol. Assess. 2018, 22, 1–60. [Google Scholar] [CrossRef]
- Mara, M.A.; Good, M.; Weitkamp, J.-H. Innate and adaptive immunity in necrotizing enterocolitis. Semin. Fetal Neonatal Med. 2018, 23, 394–399. [Google Scholar] [CrossRef]
- McElroy, S.J.; Prince, L.S.; Weitkamp, J.-H.; Reese, J.; Slaughter, J.C.; Polk, D.B. Tumor necrosis factor receptor 1-dependent depletion of mucus in immature small intestine: A potential role in neonatal necrotizing enterocolitis. Am. J. Physiol. Gastrointest. Liver Physiol. 2011, 301, G656–G666. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Sherman, M.P.; Prince, L.S.; Bader, D.; Weitkamp, J.-H.; Slaughter, J.C.; McElroy, S.J. Paneth cell ablation in the presence of Klebsiella pneumoniae induces necrotizing enterocolitis (NEC)-like injury in the small intestine of immature mice. Dis. Model. Mech. 2012, 5, 522–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lueschow, S.R.; Stumphy, J.; Gong, H.; Kern, S.L.; Elgin, T.G.; Underwood, M.A.; Kalanetra, K.M.; Mills, D.A.; Wong, M.H.; Meyerholz, D.K.; et al. Loss of murine Paneth cell function alters the immature intestinal microbiome and mimics changes seen in neonatal necrotizing enterocolitis. PLoS ONE 2018, 13, e0204967. [Google Scholar] [CrossRef] [PubMed]
- Aziz, M.M.; Ishihara, S.; Mishima, Y.; Oshima, N.; Moriyama, I.; Yuki, T.; Kadowaki, Y.; Rumi, M.A.K.; Amano, Y.; Kinoshita, Y. MFG-E8 attenuates intestinal inflammation in murine experimental colitis by modulating osteopontin-dependent alphavbeta3 integrin signaling. J. Immunol. 2009, 182, 7222–7232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, H.; Lei, Y.; He, X.; Liu, D.; He, Z. Role of lactadherin in intestinal barrier integrity in experimental neonatal necrotizing enterocolitis. J. Cell. Biochem. 2019, 120, 19509–19517. [Google Scholar] [CrossRef]
- Nair, R.R.; Warner, B.B.; Warner, B.W. Role of epidermal growth factor and other growth factors in the prevention of necrotizing enterocolitis. Semin. Perinatol. 2008, 32, 107–113. [Google Scholar] [CrossRef]
- Warner, B.B.; Ryan, A.L.; Seeger, K.; Leonard, A.C.; Erwin, C.R.; Warner, B.W. Ontogeny of salivary epidermal growth factor and necrotizing enterocolitis. J. Pediatr. 2007, 150, 358–363. [Google Scholar] [CrossRef]
- Clark, J.A.; Doelle, S.M.; Halpern, M.D.; Saunders, T.A.; Holubec, H.; Dvorak, K.; Boitano, S.A.; Dvorak, B. Intestinal barrier failure during experimental necrotizing enterocolitis: Protective effect of EGF treatment. Am. J. Physiol. Liver Physiol. 2006, 291, G938–G949. [Google Scholar] [CrossRef]
- Knott, A.W.; Juno, R.J.; Jarboe, M.D.; Zhang, Y.; Profitt, S.A.; Thoerner, J.C.; Erwin, C.R.; Warner, B.W. EGF receptor signaling affects bcl-2 family gene expression and apoptosis after massive small bowel resection. J. Pediatr. Surg. 2003, 38, 875–880. [Google Scholar] [CrossRef]
- Clark, J.A.; Lane, R.H.; Maclennan, N.K.; Holubec, H.; Dvorakova, K.; Halpern, M.D.; Williams, C.S.; Payne, C.M.; Dvorak, B. Epidermal growth factor reduces intestinal apoptosis in an experimental model of necrotizing enterocolitis. Am. J. Physiol. Gastrointest. Liver Physiol. 2005, 288, G755–G762. [Google Scholar] [CrossRef] [Green Version]
- Michalsky, M.P.; Lara-Marquez, M.; Chun, L.; Besner, G.E. Heparin-binding EGF-like growth factor is present in human amniotic fluid and breast milk. J. Pediatr. Surg. 2002, 37, 1–6. [Google Scholar] [CrossRef]
- Yang, J.; Su, Y.; Zhou, Y.; Besner, G.E. Heparin-binding EGF-like growth factor (HB-EGF) therapy for intestinal injury: Application and future prospects. Pathophysiology 2014, 21, 95–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, J.; El-Assal, O.N.; Besner, G.E. Heparin-binding EGF-like growth factor (HB-EGF) and necrotizing enterocolitis. Semin. Pediatr. Surg. 2005, 14, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Dvorak, B.; Khailova, L.; Clark, J.A.; Hosseini, D.M.; Arganbright, K.M.; Reynolds, C.A.; Halpern, M.D. Comparison of epidermal growth factor and heparin-binding epidermal growth factor-like growth factor for prevention of experimental necrotizing enterocolitis. J. Pediatr. Gastroenterol. Nutr. 2008, 47, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Sitarik, A.R.; Bobbitt, K.R.; Havstad, S.L.; Fujimura, K.E.; Levin, A.M.; Zoratti, E.M.; Kim, H.; Woodcroft, K.J.; Wegienka, G.; Ownby, D.R.; et al. Breast milk transforming growth factor β is associated with neonatal gut microbial composition. J. Pediatr. Gastroenterol. Nutr. 2017, 65, e60–e67. [Google Scholar] [CrossRef]
- Namachivayam, K.; Coffing, H.P.; Sankaranarayanan, N.V.; Jin, Y.; MohanKumar, K.; Frost, B.L.; Blanco, C.L.; Patel, A.L.; Meier, P.P.; Garzon, S.A.; et al. Transforming growth factor-β2 is sequestered in preterm human milk by chondroitin sulfate proteoglycans. Am. J. Physiol. Gastrointest. Liver Physiol. 2015, 309, G171–G180. [Google Scholar] [CrossRef] [Green Version]
- Maheshwari, A.; Kelly, D.R.; Nicola, T.; Ambalavanan, N.; Jain, S.K.; Murphy-Ullrich, J.; Athar, M.; Shimamura, M.; Bhandari, V.; Aprahamian, C.; et al. TGF-β2 suppresses macrophage cytokine production and mucosal inflammatory responses in the developing intestine. Gastroenterology 2011, 140, 242–253. [Google Scholar] [CrossRef] [Green Version]
- Bode, L. Human milk oligosaccharides in the prevention of necrotizing enterocolitis: A journey from in vitro and in vivo models to mother-infant cohort studies. Front. Pediatr. 2018, 6, 385. [Google Scholar] [CrossRef]
- Moukarzel, S.; Bode, L. Human milk oligosaccharides and the preterm infant: A journey in sickness and in health. Clin. Perinatol. 2017, 44, 193–207. [Google Scholar] [CrossRef]
- Holscher, H.D.; Bode, L.; Tappenden, K.A. Human milk oligosaccharides influence intestinal epithelial cell maturation in vitro. J. Pediatr. Gastroenterol. Nutr. 2017, 64, 296–301. [Google Scholar] [CrossRef]
- Autran, C.A.; Kellman, B.P.; Kim, J.H.; Asztalos, E.; Blood, A.B.; Spence, E.C.H.; Patel, A.L.; Hou, J.; Lewis, N.E.; Bode, L. Human milk oligosaccharide composition predicts risk of necrotising enterocolitis in preterm infants. Gut 2018, 67, 1064–1070. [Google Scholar] [CrossRef]
- Rudloff, S.; Kuntz, S.; Ostenfeldt Rasmussen, S.; Roggenbuck, M.; Sprenger, N.; Kunz, C.; Sangild, P.T.; Brandt Bering, S. Metabolism of milk oligosaccharides in preterm pigs sensitive to necrotizing enterocolitis. Front. Nutr. 2019, 6, 23. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, S.O.; Martin, L.; Østergaard, M.V.; Rudloff, S.; Roggenbuck, M.; Nguyen, D.N.; Sangild, P.T.; Bering, S.B. Human milk oligosaccharide effects on intestinal function and inflammation after preterm birth in pigs. J. Nutr. Biochem. 2017, 40, 141–154. [Google Scholar] [CrossRef] [PubMed]
- Jantscher-Krenn, E.; Zherebtsov, M.; Nissan, C.; Goth, K.; Guner, Y.S.; Naidu, N.; Choudhury, B.; Grishin, A.V.; Ford, H.R.; Bode, L. The human milk oligosaccharide disialyllacto-N-tetraose prevents necrotising enterocolitis in neonatal rats. Gut 2012, 61, 1417–1425. [Google Scholar] [CrossRef] [PubMed]
- Good, M.; Sodhi, C.P.; Yamaguchi, Y.; Jia, H.; Lu, P.; Fulton, W.B.; Martin, L.Y.; Prindle, T.; Nino, D.F.; Zhou, Q.; et al. The human milk oligosaccharide 2’-fucosyllactose attenuates the severity of experimental necrotising enterocolitis by enhancing mesenteric perfusion in the neonatal intestine. Br. J. Nutr. 2016, 116, 1175–1187. [Google Scholar] [CrossRef] [Green Version]
- Agostoni, C.; Carratù, B.; Boniglia, C.; Lammardo, A.M.; Riva, E.; Sanzini, E. Free glutamine and glutamic acid increase in human milk through a three-month lactation period. J. Pediatr. Gastroenterol. Nutr. 2000, 31, 508–512. [Google Scholar] [CrossRef]
- Baldeón, M.E.; Zertuche, F.; Flores, N.; Fornasini, M. Free amino acid content in human milk is associated with infant gender and weight gain during the first four months of lactation. Nutrients 2019, 11, 2239. [Google Scholar] [CrossRef] [Green Version]
- Larnkjær, A.; Bruun, S.; Pedersen, D.; Zachariassen, G.; Barkholt, V.; Agostoni, C.; Mlgaard, C.; Husby, S.; Michaelsen, K.F. Free amino acids in human milk and associations with maternal anthropometry and infant growth. J. Pediatr. Gastroenterol. Nutr. 2016, 63, 374–378. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.-H.; Kim, H. The roles of glutamine in the intestine and its implication in intestinal diseases. Int. J. Mol. Sci. 2017, 18, 1051. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Wu, G.; Zhou, Z.; Dai, Z.; Sun, Y.; Ji, Y.; Li, W.; Wang, W.; Liu, C.; Han, F.; et al. Glutamine and intestinal barrier function. Amino Acids 2015, 47, 2143–2154. [Google Scholar] [CrossRef]
- Becker, R.M.; Wu, G.; Galanko, J.A.; Chen, W.; Maynor, A.R.; Bose, C.L.; Rhoads, J.M. Reduced serum amino acid concentrations in infants with necrotizing enterocolitis. J. Pediatr. 2000, 137, 785–793. [Google Scholar] [CrossRef]
- Zhou, W.; Li, W.; Zheng, X.-H.; Rong, X.; Huang, L.-G. Glutamine downregulates TLR-2 and TLR-4 expression and protects intestinal tract in preterm neonatal rats with necrotizing enterocolitis. J. Pediatr. Surg. 2014, 49, 1057–1063. [Google Scholar] [CrossRef] [PubMed]
- Sodhi, C.P.; Neal, M.D.; Siggers, R.; Sho, S.; Ma, C.; Branca, M.F.; Prindle, T.; Russo, A.M.; Afrazi, A.; Good, M.; et al. Intestinal epithelial Toll-like receptor 4 regulates goblet cell development and is required for necrotizing enterocolitis in mice. Gastroenterology 2012, 143, 708–718.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Good, M.; Sodhi, C.P.; Egan, C.E.; Afrazi, A.; Jia, H.; Yamaguchi, Y.; Lu, P.; Branca, M.F.; Ma, C.; Prindle, T.; et al. Breast milk protects against the development of necrotizing enterocolitis through inhibition of Toll-like receptor 4 in the intestinal epithelium via activation of the epidermal growth factor receptor. Mucosal Immunol. 2015, 8, 1166–1179. [Google Scholar] [CrossRef] [PubMed]
- Sodhi, C.P.; Shi, X.-H.; Richardson, W.M.; Grant, Z.S.; Shapiro, R.A.; Prindle, T.; Branca, M.; Russo, A.; Gribar, S.C.; Ma, C.; et al. Toll-like receptor-4 inhibits enterocyte proliferation via impaired beta-catenin signaling in necrotizing enterocolitis. Gastroenterology 2010, 138, 185–196. [Google Scholar] [CrossRef] [Green Version]
- El-Shimi, M.S.; Awad, H.A.; Abdelwahed, M.A.; Mohamed, M.H.; Khafagy, S.M.; Saleh, G. Enteral L-arginine and glutamine supplementation for prevention of NEC in preterm neonates. Int. J. Pediatr. 2015, 2015, 856091. [Google Scholar] [CrossRef] [Green Version]
- Brown, J.; Moe-Byrne, T.; McGuire, W. Glutamine supplementation for young infants with severe gastrointestinal disease. Cochrane Database Syst. Rev. 2007, CD005947. [Google Scholar] [CrossRef] [Green Version]
- Moe-Byrne, T.; Brown, J.V.E.; McGuire, W. Glutamine supplementation to prevent morbidity and mortality in preterm infants. Cochrane Database Syst. Rev. 2016, 4, CD001457. [Google Scholar]
- Zhou, L.; Yoshimura, Y.; Huang, Y.; Suzuki, R.; Yokoyama, M.; Okabe, M.; Shimamura, M. Two independent pathways of maternal cell transmission to offspring: Through placenta during pregnancy and by breast-feeding after birth. Immunology 2000, 101, 570–580. [Google Scholar] [CrossRef]
- Lewis, E.D.; Richard, C.; Larsen, B.M.; Field, C.J. The importance of human milk for immunity in preterm infants. Clin. Perinatol. 2017, 44, 23–47. [Google Scholar] [CrossRef]
- Trend, S.; de Jong, E.; Lloyd, M.L.; Kok, C.H.; Richmond, P.; Doherty, D.A.; Simmer, K.; Kakulas, F.; Strunk, T.; Currie, A. Leukocyte populations in human preterm and term breast milk identified by multicolour flow cytometry. PLoS ONE 2015, 10, e0135580. [Google Scholar] [CrossRef] [Green Version]
- Cabinian, A.; Sinsimer, D.; Tang, M.; Zumba, O.; Mehta, H.; Toma, A.; Sant’Angelo, D.; Laouar, Y.; Laouar, A. Transfer of maternal immune cells by breastfeeding: Maternal cytotoxic T lymphocytes present in breast milk localize in the peyer’s patches of the nursed infant. PLoS ONE 2016, 11, e0156762. [Google Scholar] [CrossRef] [PubMed]
- Riskin, A.; Almog, M.; Peri, R.; Halasz, K.; Srugo, I.; Kessel, A. Changes in immunomodulatory constituents of human milk in response to active infection in the nursing infant. Pediatr. Res. 2012, 71, 220–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ustundag, B.; Yilmaz, E.; Dogan, Y.; Akarsu, S.; Canatan, H.; Halifeoglu, I.; Cikim, G.; Aygun, A.D. Levels of cytokines (IL-1beta, IL-2, IL-6, IL-8, TNF-alpha) and trace elements (Zn, Cu) in breast milk from mothers of preterm and term infants. Mediators Inflamm. 2005, 2005, 331–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minekawa, R.; Takeda, T.; Sakata, M.; Hayashi, M.; Isobe, A.; Yamamoto, T.; Tasaka, K.; Murata, Y. Human breast milk suppresses the transcriptional regulation of IL-1beta-induced NF-kappaB signaling in human intestinal cells. Am. J. Physiol. Cell Physiol. 2004, 287, C1404–C1411. [Google Scholar] [CrossRef] [Green Version]
- Bryan, D.-L.; Forsyth, K.D.; Gibson, R.A.; Hawkes, J.S. Interleukin-2 in human milk: A potential modulator of lymphocyte development in the breastfed infant. Cytokine 2006, 33, 289–293. [Google Scholar] [CrossRef]
- Hassan, J.; Reen, D.J. Reduced primary antigen-specific T-cell precursor frequencies in neonates is associated with deficient interleukin-2 production. Immunology 1996, 87, 604–608. [Google Scholar] [CrossRef]
- Saito, S.; Maruyama, M.; Kato, Y.; Moriyama, I.; Ichijo, M. Detection of IL-6 in human milk and its involvement in IgA production. J. Reprod. Immunol. 1991, 20, 267–276. [Google Scholar] [CrossRef]
- Rudloff, H.E.; Schmalstieg, F.C.; Palkowetz, K.H.; Paszkiewicz, E.J.; Goldman, A.S. Interleukin-6 in human milk. J. Reprod. Immunol. 1993, 23, 13–20. [Google Scholar] [CrossRef]
- Maheshwari, A.; Lu, W.; Lacson, A.; Barleycorn, A.A.; Nolan, S.; Christensen, R.D.; Calhoun, D.A. Effects of interleukin-8 on the developing human intestine. Cytokine 2002, 20, 256–267. [Google Scholar] [CrossRef]
- Polat, A.; Tunc, T.; Erdem, G.; Yerebasmaz, N.; Tas, A.; Beken, S.; Basbozkurt, G.; Saldir, M.; Zenciroglu, A.; Yaman, H. Interleukin-8 and its receptors in human milk from mothers of full-term and premature infants. Breastfeed. Med. 2016, 11, 247–251. [Google Scholar] [CrossRef]
- Fiorentino, D.F.; Bond, M.W.; Mosmann, T.R. Two types of mouse T helper cell. IV. Th2 clones secrete a factor that inhibits cytokine production by Th1 clones. J. Exp. Med. 1989, 170, 2081–2095. [Google Scholar] [CrossRef] [PubMed]
- Garofalo, R.; Chheda, S.; Mei, F.; Palkowetz, K.H.; Rudloff, H.E.; Schmalstieg, F.C.; Rassin, D.K.; Goldman, A.S. Interleukin-10 in human milk. Pediatr. Res. 1995, 37, 444–449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fiorentino, D.F.; Zlotnik, A.; Mosmann, T.R.; Howard, M.; O’Garra, A. IL-10 inhibits cytokine production by activated macrophages. J. Immunol. 1991, 147, 3815–3822. [Google Scholar] [PubMed]
- Jinquan, T.; Larsen, C.G.; Gesser, B.; Matsushima, K.; Thestrup-Pedersen, K. Human IL-10 is a chemoattractant for CD8+ T lymphocytes and an inhibitor of IL-8-induced CD4+ T lymphocyte migration. J. Immunol. 1993, 151, 4545–4551. [Google Scholar] [PubMed]
- Fluckiger, A.C.; Garrone, P.; Durand, I.; Galizzi, J.P.; Banchereau, J. Interleukin 10 (IL-10) upregulates functional high affinity IL-2 receptors on normal and leukemic B lymphocytes. J. Exp. Med. 1993, 178, 1473–1481. [Google Scholar] [CrossRef] [Green Version]
- Ballard, O.; Morrow, A.L. Human milk composition: Nutrients and bioactive factors. Pediatr. Clin. N. Am. 2013, 60, 49–74. [Google Scholar] [CrossRef] [Green Version]
- Beaurepaire, C.; Smyth, D.; McKay, D.M. Interferon-gamma regulation of intestinal epithelial permeability. J. Interferon Cytokine Res. 2009, 29, 133–144. [Google Scholar] [CrossRef]
- Hui, L.; Dai, Y.; Guo, Z.; Zhang, J.; Zheng, F.; Bian, X.; Wu, Z.; Jiang, Q.; Guo, M.; Ma, K.; et al. Immunoregulation effects of different γδT cells and toll-like receptor signaling pathways in neonatal necrotizing enterocolitis. Medicine (Baltimore). 2017, 96, e6077. [Google Scholar] [CrossRef]
- Yu, J.C.; Khodadadi, H.; Malik, A.; Davidson, B.; da Salles, É.S.L.; Bhatia, J.; Hale, V.L.; Baban, B. Innate immunity of neonates and infants. Front. Immunol. 2018, 9, 1759. [Google Scholar] [CrossRef]
- Buescher, E.S.; Malinowska, I. Soluble receptors and cytokine antagonists in human milk. Pediatr. Res. 1996, 40, 839–844. [Google Scholar] [CrossRef] [Green Version]
- Castellote, C.; Casillas, R.; Ramírez-Santana, C.; Pérez-Cano, F.J.; Castell, M.; Moretones, M.G.; López-Sabater, M.C.; Franch, A. Premature delivery influences the immunological composition of colostrum and transitional and mature human milk. J. Nutr. 2011, 141, 1181–1187. [Google Scholar] [CrossRef] [PubMed]
- Chokshi, N.K.; Guner, Y.S.; Hunter, C.J.; Upperman, J.S.; Grishin, A.; Ford, H.R. The role of nitric oxide in intestinal epithelial injury and restitution in neonatal necrotizing enterocolitis. Semin. Perinatol. 2008, 32, 92–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nankervis, C.A.; Giannone, P.J.; Reber, K.M. The neonatal intestinal vasculature: Contributing factors to necrotizing enterocolitis. Semin. Perinatol. 2008, 32, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Akçay, F.; Aksoy, H.; Memişoǧullari, R. Effect of breast-feeding on concentration of nitric oxide in breast milk. Ann. Clin. Biochem. 2002, 39, 68–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ford, H.; Watkins, S.; Reblock, K.; Rowe, M. The role of inflammatory cytokines and nitric oxide in the pathogenesis of necrotizing enterocolitis. J. Pediatr. Surg. 1997, 32, 275–282. [Google Scholar] [CrossRef]
- Yazji, I.; Sodhi, C.P.; Lee, E.K.; Good, M.; Egan, C.E.; Afrazi, A.; Neal, M.D.; Jia, H.; Lin, J.; Ma, C.; et al. Endothelial TLR4 activation impairs intestinal microcirculatory perfusion in necrotizing enterocolitis via eNOS-NO-nitrite signaling. Proc. Natl. Acad. Sci. USA 2013, 110, 9451–9456. [Google Scholar] [CrossRef] [Green Version]
- Bravi, F.; Wiens, F.; Decarli, A.; Dal Pont, A.; Agostoni, C.; Ferraroni, M. Impact of maternal nutrition on breast-milk composition: A systematic review. Am. J. Clin. Nutr. 2016, 104, 646–662. [Google Scholar] [CrossRef] [Green Version]
- Gay, M.C.L.; Koleva, P.T.; Slupsky, C.M.; du Toit, E.; Eggesbo, M.; Johnson, C.C.; Wegienka, G.; Shimojo, N.; Campbell, D.E.; Prescott, S.L.; et al. Worldwide variation in human milk metabolome: Indicators of breast physiology and maternal lifestyle? Nutrients 2018, 10, 1151. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Overview of the immunomodulatory components of maternal breast milk.


{kind=link}
Table 1.
Cytokines present in human breast milk and physiologic relevance to the infant.
| Cytokine | Composition in Human Milk and Significance | References |
|---|---|---|
| Interleukin (IL)-1 |
| [53,64] |
| IL-2 |
| [63,65,66] |
| IL-6 |
| [63,67,68] |
| IL-8 |
| [63,69,70] |
| IL-10 |
| [71,72,73,74,75] |
| IFN-γ |
| [76,77,78,79] |
| TNF-α |
| [63,80,81] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Nolan, L.S.; Parks, O.B.; Good, M. A Review of the Immunomodulating Components of Maternal Breast Milk and Protection Against Necrotizing Enterocolitis. Nutrients 2020, 12, 14. https://doi.org/10.3390/nu12010014
AMA Style
Nolan LS, Parks OB, Good M. A Review of the Immunomodulating Components of Maternal Breast Milk and Protection Against Necrotizing Enterocolitis. Nutrients. 2020; 12(1):14. https://doi.org/10.3390/nu12010014
Chicago/Turabian StyleNolan, Lila S., Olivia B. Parks, and Misty Good. 2020. "A Review of the Immunomodulating Components of Maternal Breast Milk and Protection Against Necrotizing Enterocolitis" Nutrients 12, no. 1: 14. https://doi.org/10.3390/nu12010014
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.