1. Introduction
Humans are inherently social creatures. They cooperate to satisfy physiological and affiliative needs in terms of care, protection, and reproduction [
1], thus safeguarding their survival from environmental hazards [
2] and increasing their sense of inclusiveness [
3].
From the first years of life, infants are exposed to a stimulating environment that strengthens their social bonds with parents and family members [
4]. During infancy and childhood, the individual lays the basis for a complex pattern of exchange and engagement through his/her interaction with relatives and peers [
5]. The distinctive pattern in early attachment with caregivers can be stable across human development and influence adult attachment with partners [
6,
7]. Within the interaction between caregivers and offspring [
8], high quality parental bonding enhances children’s self-efficacy [
9], as well as social and emotional communication [
10], while reducing psychological [
11] and physiological [
12,
13] stress. Parental bonding represents the core causal factor of the main developmental stages, which remain open to modulation due to experience and exposure to further environmental events [
14].
Currently, both digital and physical spaces can forge social bonds and become the theaters of online and offline interactions that affect relationships among people. The adoption of technological devices allows children to explore social media platforms and online worlds [
15]. Hence, the growing social skills in virtual and real environments shape adolescents’ [
16,
17,
18] and adults’ online and offline behaviors [
19,
20,
21]. In particular, the ubiquitous usage of Social Network Sites (SNSs) has created a virtual environment where social interactions can happen anytime and anywhere [
22,
23,
24,
25,
26]. Among the different platforms, Instagram, one of the most popular sites for youths [
27], focuses on photo-sharing and visual content. It is mainly characterized by various online social behaviors such as scrolling through followings’ contents, watching stories, publishing posts, tagging followers, commenting on photos, and chatting privately [
28].
With the advent of Instagram, kids are early participants of social media. Parents become prey to “sharenting”, the phenomenon that describes the tendency to exhibit pictures of their children online [
29]. Interestingly, recent findings have highlighted a disagreement regarding the positive and negative outcomes of online social interactions on SNSs. For instance, the frequency of Facebook and Instagram usage, as well as high rates in reported self-attractiveness [
30] are associated not only with a higher self-esteem [
31], but also with depressive symptoms [
32]. Furthermore, Instagram improves happiness and decreases solitude with the intimacy offered by the images [
33]. At the same time, an increased number of Instagram posting activities, combined with individual body dissatisfaction, increases the probability of engaging in negative romantic relationships [
34]. The reason for Instagram use was also traced to a high need for interpersonal interaction and dominant traits in narcissism [
35,
36]. The same parents share their “family snapshots” to prove that the “merry family ideal” and the “cute child ideal” have come true [
37]. Although several papers investigated how parental factors are associated with user activity on Facebook [
38,
39,
40] and Instagram [
29,
37,
41], there is a lack of research focused on the interplay between Instagram activity and early attachment with parents. This gap is even more evident if the potential relationship between attachment and genetic factors is considered. Only a few studies recently probed the genetic influence on the frequency of social media use [
42,
43].
The evolutionary tendency to be social can be modulated by genetic factors, which, in turn, are regulated by the environmental actions across the human lifespan, thus conferring nuanced levels of sensitivity to the experiences. Based on the model of the plasticity of genes [
44,
45], it is not only the protective factor versus the risk factor of genes that determines human behavior. Indeed, one must also take into consideration the quality of environmental factors that interact with allelic expression in shaping physiological responses and then behavioral patterns that foster environmental adaptation [
46].
According to the susceptibility hypothesis or sensitivity hypothesis [
47], alleles (i.e., G or A) of a given genetic region (i.e., oxytocin receptor gene rs2254298) are associated with different degrees of sensitivity to the environmental effects (i.e., quality of early parental care) [
48]. Here, a genetic risk factor makes the individual genetically sensitive or susceptible to life events and individual experience [
49,
50].
In the context of sociability, people with high genetic sensitivity will exhibit a more adaptive social behavior when exposed to a positive environment (i.e., a warm relationship with parents) [
51,
52]. However, they will display less adaptive social behavior if they go through negative events (i.e., the loss of a parent, child abuse, or maltreatment) [
53,
54]. Conversely, low genetic sensitivity and vulnerability will make someone more resistant to the effects of early traumatic episodes (i.e., parental separation or abandonment), as well as beneficial events (i.e., competent caregiving), generating a decreased social response [
55].
Within this debate, multiple studies have attributed a key role in the biological explanation of social behavior to the hormone oxytocin [
56,
57,
58]. Specifically, rs53576 and rs2254298 polymorphisms, encoded by the Oxytocin receptor gene (OXTr), correlate with social behaviors, social cognition, and empathy [
59,
60]. For each region, two allelic structures have been observed to play a role as determinants in social development: guanine (G) to adenine (A) substitution shows greater sensitivity to the environment and influences responses to stressful life events [
61]. However, from the previous results, it is not possible to unequivocally establish which variation—G or A—is more associated with less adaptive social responses.
Concerning rs2254298, a history of paternal overprotection was found to moderate the heart rate responses to socially distressing stimuli (increased for A-carriers, but decreased for G/G homozygotes) [
62]. A-carriers also showed lower empathetic levels than G-carriers regardless of parental warmth [
60].
As for rs53576, individuals with the G variation show a variety of favorable features when compared to A-carriers, such as higher levels of trust [
63], dispositional empathy [
64], greater sympathetic response to stressors [
50], and more sensitivity to social cues [
65] and social interactions [
66]. Moreover, social support before a distressing task was observed to reduce cortisol response in G-carriers, but not in A/A homozygotes [
67].
Taken together, these scientific contributions point out that genetic expression, combined with early environmental exposure, contribute to the shaping of adult sociability. However, the role played by specific genetic predispositions (i.e., the ones related to early environment exposure during infant-parent interaction) on the online social relationships of adults is considerably underexplored.
This study investigates how OXTr and caregivers’ propensities during childhood interact in modulating adult online relationships on Instagram (see
Appendix A). Specifically, two OXTr Single-Nucleotide Polymorphisms (OXTr SNPs: rs2254298 and rs53576) for the genetic component and the Parental Bonding Instrument as the assessment for the parent-child recalled bonding were considered. Likewise, the Instagram number of (i) followed users (here called “followings”), (ii) published posts, (iii) followers, and (iv) a further combined index, called the “Social Desirability Index” (SDI), were selected as the main social media variables.
This goal was addressed with the formulation of a combined directional hypothesis to evaluate both correlations and mean level similarities and differences between conditions. In favor of the sensitivity hypotheses, for each Instagram variable, an interaction effect between the genetic component and the parental attachment scores, independent of gender, was hypothesized. More specifically, adult Instagram users with a genetic risk factor (OXTr rs2254298 A-carriers, OXTr rs53576 G-carriers) and who were exposed to a beneficial and positive early relationship with their parents (high parental care, low parental overprotection) would show increased online social activity (higher number of posts and followings) compared to less vulnerable genetic carriers (OXTr rs2254298 G/G homozygotes, OXTr rs53576 A/A homozygotes). Conversely, Instagram users with a genetic risk factor and who were exposed to an adverse and negative early relationship with parents (low parental care, high parental overprotection) would show decreased online social activity as described by a lower number of posts and followings compared to less vulnerable genetic carriers.
4. Discussion
This study investigated how alleles in OXTr rs2254298 and rs53576 interact differently with parental care and overprotection during childhood in explaining Instagram social behavior. Two genotype*environment interactions (OXTr rs2254298 SNP * parental bonding in childhood; OXTr rs53576 SNP * parental bonding in childhood) on the number of Instagram followings and posts were hypothesized.
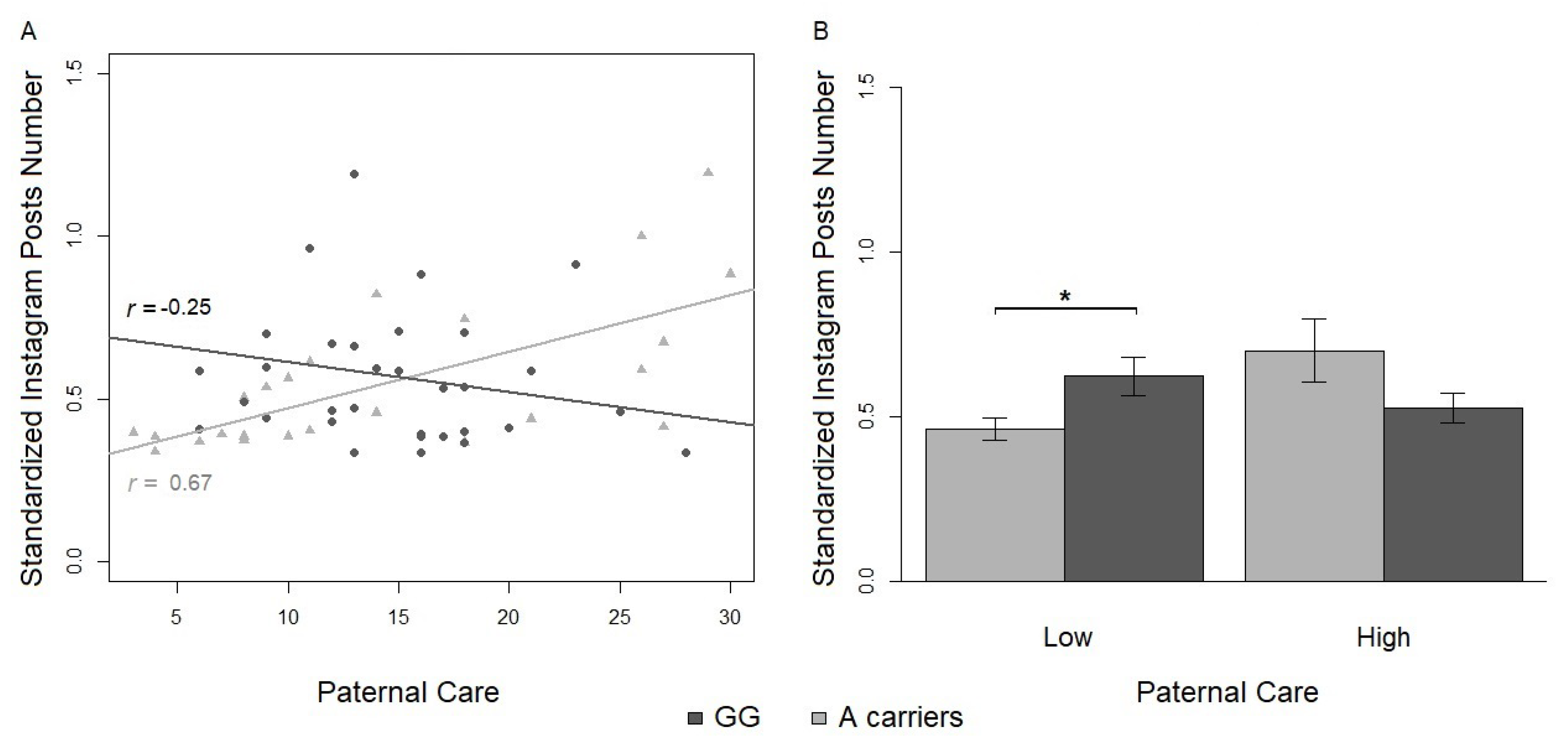
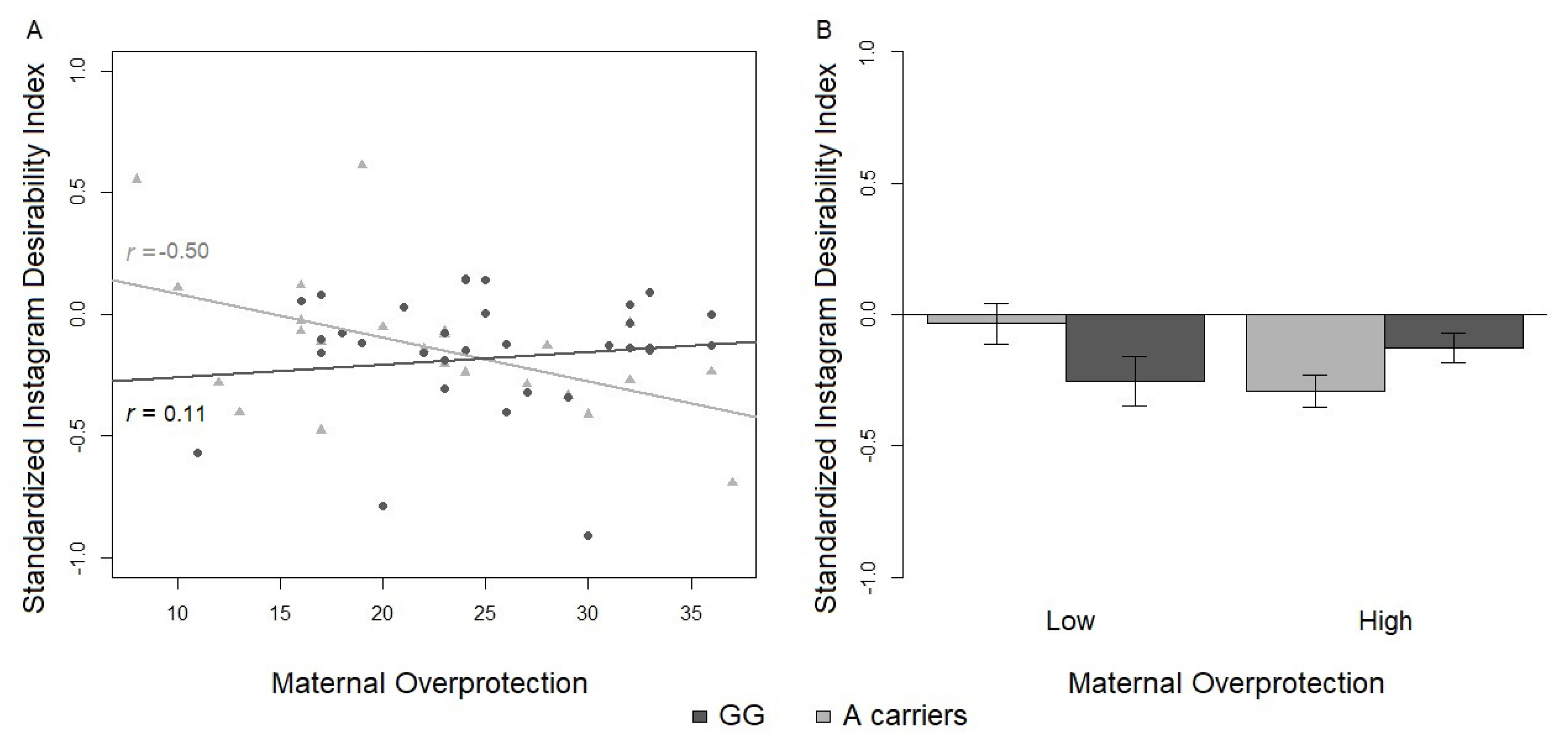
In line with the hypotheses in this study, adult Instagram users with a genetic risk factor (OXTr rs2254298 G/A or A/A genotype) show differential Instagram social activity according to their own early experience with parents. In particular, A-carriers, when exposed to a less optimal early environment (represented by low scores in parental care), showed a lower number of Instagram posts compared to less vulnerable genetic carriers (G/G homozygotes). Interestingly, A-carriers with a reported history of maternal overprotection also showed a decreasing trend in the Instagram SDI, whereas those with low scores in maternal overprotection showed an increasing trend in the same index. Overall, A-carriers who experienced negative patterns of interactions with their caregiver in childhood, as indicated by low paternal care and high maternal overprotection, exhibited weakened social responses on Instagram. From an analytical view, both interaction effects were found to survive the magnitude of the Bonferroni correction, which was differentially applied for the predicted (i.e., on the number of Instagram posts) and exploratory analysis (i.e., on the Instagram SDI).
To the authors’ knowledge, this is the first study that analyzes Instagram activity from a gene-environment perspective. Specifically, the present research examines Instagram behavior, a novel area to be explored, on a non-Western sample, which is usually under-represented in psychological studies [
90]. Furthermore, these results corroborate the literature, which highlights the interaction effects between genetic predispositions and early social behaviors on human development. The moderating impact of genetics on environmental effects over Instagram social activity underpins the sensitivity hypotheses [
91]. Within this framework, researchers have asserted that humans less susceptible to the environment (OXTr rs2254298 G/G homozygotes) will be less affected by stressful and negative conditions, but also by calming and positive events [
44,
61]. In contrast, in terms of conditional adaptation, humans with higher biological innate environmental susceptibility (OXTr rs2254298 A-carriers) display two dissociated behavioral patterns in relation to the quality of their early caring experiences and the consequent psychosocial outcomes [
46,
56,
92,
93]: (a) if exposed to a maladaptive environment and met with strenuous adversities, these individuals would show poorer sociable attitudes towards conspecifics compared to less susceptible individuals; (b) if exposed to an adaptive environment and functional experiences, these individuals would show higher functional social characteristics in the course of development and would cope better with stressful events compared to less sensitive individuals [
45,
48,
94,
95].
In connection with the present findings, the G allele predisposes users to be less vulnerable, but also less plastic to the environment, thus displaying a relatively fixed Instagram social pattern less associated with the quality of early parental caregiving; whereas the A allele increases the vulnerability to the environment and moderates online social behavior: (a) individuals with low paternal care are less social users (in terms of low number of posts) than G/G homozygotes on Instagram; (b) individuals with low maternal overprotection show an opposite trend of general sociability (in terms of high SDI) compared to G/G homozygotes on Instagram. However, it is important to specify the parameters from which a differential level of sociability as a function of the two cross gene*environment interactions can be inferred.
Firstly, A-carriers with low paternal care posted less than G/G homozygotes. A plausible argument is that lower paternal care could have determined decreased abilities in coping with Instagram’s overstimulating and socially demanding environment. In turn, less adaptive online behaviors could be explained by users’: (a) reluctance to be a proactive agent of the social network; (b) reluctance to be exposed to others’ judgment or influence [
81,
96]. An alternative explanation could be that these users are not interested in self-promotion or the development of social ties by posting [
97].
Secondly, A-carriers with high levels of maternal overprotection showed a decreasing trend in the Instagram SDI, whereas the opposite pattern was observed in A-carriers with lower levels of maternal overprotection. The SDI was calculated to estimate the asymmetry between the number of followers and followings for each Instagram account. Here, a history of high maternal overprotection, potentially linked to the repression of child’s emotional expressions and autonomy [
98], could shape the structure of the Instagram user’s social network: the higher the SDI, the higher the number of followers at the expense of the number of followings, the higher the recognition that each user receives from other Instagrammers. This being said, a higher grade of social desirability could be more likely achieved with increased self-management [
99], more intense online activity, and the ability to attend to followers’ requests or expectations [
28], thus all attitudes that could be undermined by high levels of maternal overprotection experienced in childhood [
100,
101]. This index could be even more useful in clustering different groups of users and distributing the scoring along a continuum from a minimum to a maximum number of followers over followings. Users who are not skilled with Instagram services will show a limited number of followers, while more competent users with good quality content will get higher follow back rates. Interestingly, Instagram posting activity and the SDI results could be strongly linked parameters since the former could characterize the first means to enhance the latter.
To conclude, Instagram users were affected by their reported paternal bonding, as well as maternal bonding. The detection of interaction effects that included one dimension for each parent provides a more balanced representation of the potential contribution of the parents in caregiving behavior.
From an educational perspective, a relationship promoting dialogue between parents and kids could represent a valuable resource in guiding social behavior towards peers [
102] and supervising Internet and social media usage [
103]. A trusting, but not excessively controlling presence of the parents may allow increasing the autonomy of the child in managing social media content [
104]. A moderating action of parents in the administration of online life (i.e., defining a maximum duration of use, watching digital content with parents, discussing and clarifying the visualized content with parents) can train their child’s ability to recognize the potential risks of the web [
105] and reduce the risks of developing addiction associated with the problematic use of the Internet and online services (i.e., Internet use disorder, Internet gaming disorder, Hikikomori syndrome) [
106,
107,
108], such as social media [
109,
110].
Nevertheless, no significant interaction effects between OXTr rs53576 and parental bonding in childhood on Instagram social behaviors were detected. This result prompted us to elaborate on the following reasons. At a pragmatic level, different from rs2254298, the distribution in the sample for rs53576 A/A homozygous and G-carriers did not find total correspondence with the range of averaged distributions of these genotypes in the general Asiatic population. There could be indeed a remarkable variability in the alleles’ frequency in the Asiatic population, for example between East and South Asia (1000 Genomes project, BioSamples: SAMN07486027-SAMN07486024, dbSNP (Short Genetic Variations), 2017). Concerning this, it is arduous to ascertain the genetic distribution in a country like Singapore. At a more theoretical level, although there are several pieces of evidence of correlations between rs53576 polymorphism and pro-social tendencies and empathy [
65,
111], rs53576 could be more related to other explicit forms of sociality than online sociality itself. This statement could find support from two meta-analyses [
66,
112], which found a lack of association between rs53576 and general sociality. These considerations, taken together, could point to further investigation on the specific functionality of the single-nucleotide polymorphism in the region of rs53576.
Good caregiving practices could boost social approach, even in SNSs. In light of these considerations, oxytocin receptor gene polymorphisms (rs2254298) and early caregiving behaviors contribute to the modulation of Instagram user behavior. These findings shed light on a specific side of social behavior: an online social marker of the interaction between genetic and environmental factors.
5. Limitations and Future Directions
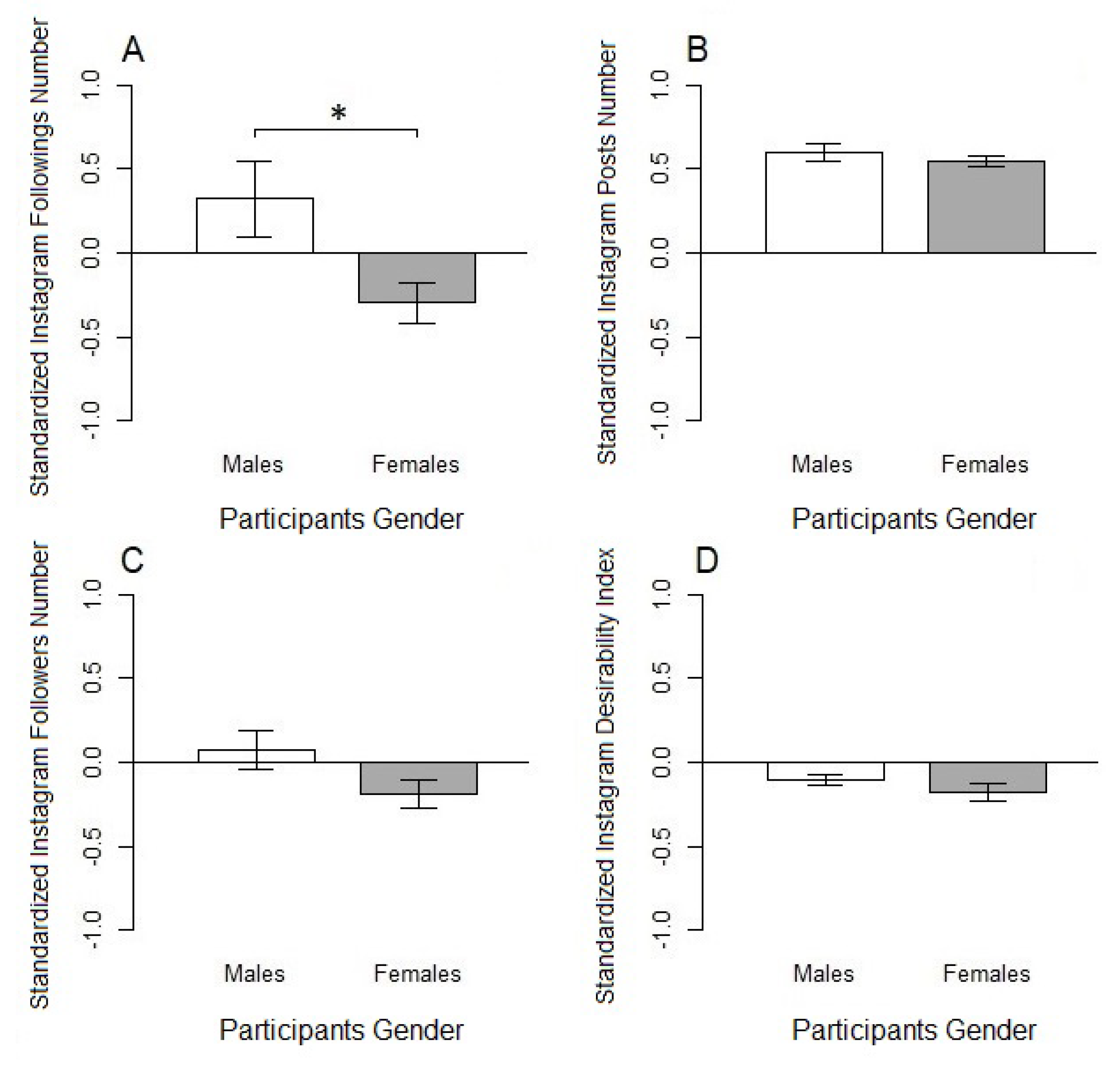
The current study has several limitations. First of all, the sample size was determined by study constraints and had a prevalence of the female gender. In the last decade, genetic association studies with limited sample sizes have become the target of increasing concerns related to the small variance explained in variables by one single-nucleotide polymorphisms (i.e., OXTr rs53576) [
113,
114]. However, given the difficulty of unraveling the nature of a given psychological construct (i.e., online social behavior) in a gene-environment perspective, the same experts of the fields cannot provide a definitive and unique solution to this implicit limitation [
113]. For instance, the only evidence that oxytocin is a neuropeptide highly involved in a variety of functions [
115] and expressed in several brain regions could exponentially boost the probability of detecting a significant effect [
116]. A large number of loci could explain a given phenotype [
117]. Here, only the genetic candidates OXTr rs2254298 and rs53576 were selected with care based on their well-documented functional properties [
67], the hypothesis formulated for this study, and the results from previous works in the field [
50,
62]. From a statistical viewpoint, Cohen’s rule of interpreting the magnitude of a significant result [
118] gave evidence that the significant effects (i.e., the interaction between paternal care and OXTr rs2254298 over Instagram posts number; the interaction between maternal overprotection and OXTr rs2254298 over the Social Desirability Index) were medium. The effect sizes were also consistent with the post-hoc power estimation. Other studies of behavioral genetics found comparable post-hoc statistical power in a small sample size [
50,
62]. Despite the efforts in data collection and genotyping, the current results should be interpreted with caution in light of the current debate on genetic association studies. Although it was not possible to maximize the sample size, a conservative approach to data treatment was adopted, and the results were in line with the expected patterns and defined by a reliable statistical magnitude. This study, in an exploratory way, deals with gene-environment interactions applied to new and recent variables from social media (i.e., Instagram social behavior), thus paving the way for further studies on this novel phenomenon.
Concerning the second limitation, no significant interaction effects were found for OXTr rs53576, presumably due to an unbalanced distribution in allele frequency within the sample. Further investigation could focus more on the role of OXTr rs53576 as a potential moderator of online social dispositions on SNSs such as Instagram. Third, the self-reported PBI questionnaire provides a retrospective measure of the individual self-perception of attachment, which could lead to influential biases. Alternative paradigms could include: (a) observational techniques as a more direct measure of caregiving patterns in an ecological context; (b) a longitudinal approach able to collect information on parental practices from childhood to adolescence. Fourth, four a priori objective parameters that were able to provide a reliable measure of each participant’s Instagram behavior were selected. Subsequent research could adopt different variables or indexes to explore the variegated world of Instagram. Overall, future studies should consider advanced interactions between new genetic factors (i.e., genetic polymorphisms of serotonin, dopamine) and environmental factors (i.e., membership within peer groups, socioeconomic status), but also hormonal components (i.e., menstrual stages in women, testosterone levels in men) and social characteristics (i.e., adult attachment with partners, quality of adult relationships) and individual traits (i.e., personality, education). All together, these variables could grasp common and specific mechanisms that characterize offline versus online social behavior.

 ,
, 




{kind=link}
{kind=link}
{kind=link}