Children’s DAT1 Polymorphism Moderates the Relationship Between Parents’ Psychological Profiles, Children’s DAT Methylation, and Their Emotional/Behavioral Functioning in a Normative Sample

 ,
,  and
and
Abstract
:1. Introduction
1.1. Dopamine Transporter Gene
1.2. Gene x Environment Interactions for DAT1
1.3. Beyond GxE: The Role of DAT1 Methylation
1.4. The Present Study
- (1)
- The possible predictive effect of parental psychological profile on children’s DAT methylation;
- (2)
- The possible predictive effect of the influence of children’s DAT methylation status on their own emotional/behavioral functioning;
- (3)
- The possible relationship between parents’ psychological profile and children’s emotional/behavioral functioning.
2. Materials and Methods
2.1. Procedure and Sample
Procedure for Biological Sampling
2.2. Instruments
2.2.1. Assessment of Parents’ Psychopathological Symptoms
2.2.2. Assessment of Parents’ Emotional/Behavioral Functioning
2.2.3. Assessment of Children’s Emotional/Behavioral Functioning
2.2.4. DNA Isolation and Genotyping
2.2.5. Analysis of DNA Methylation
2.3. Statistical Analysis
3. Results
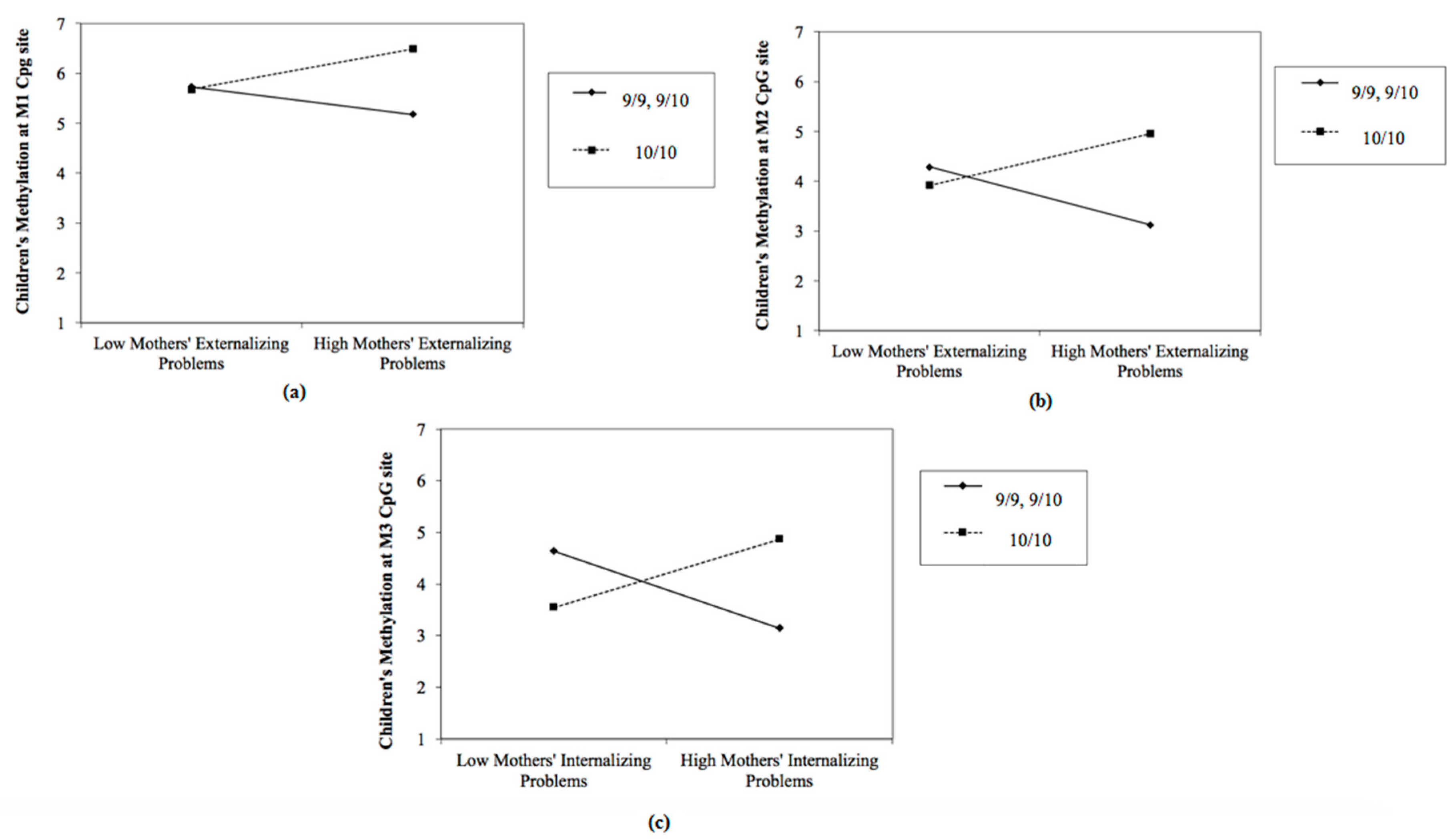
3.1. The Predictive Effect of Parents’ Psychological Profile on Children’s DAT Methylation, Moderated by Children’s DAT1 Genotype
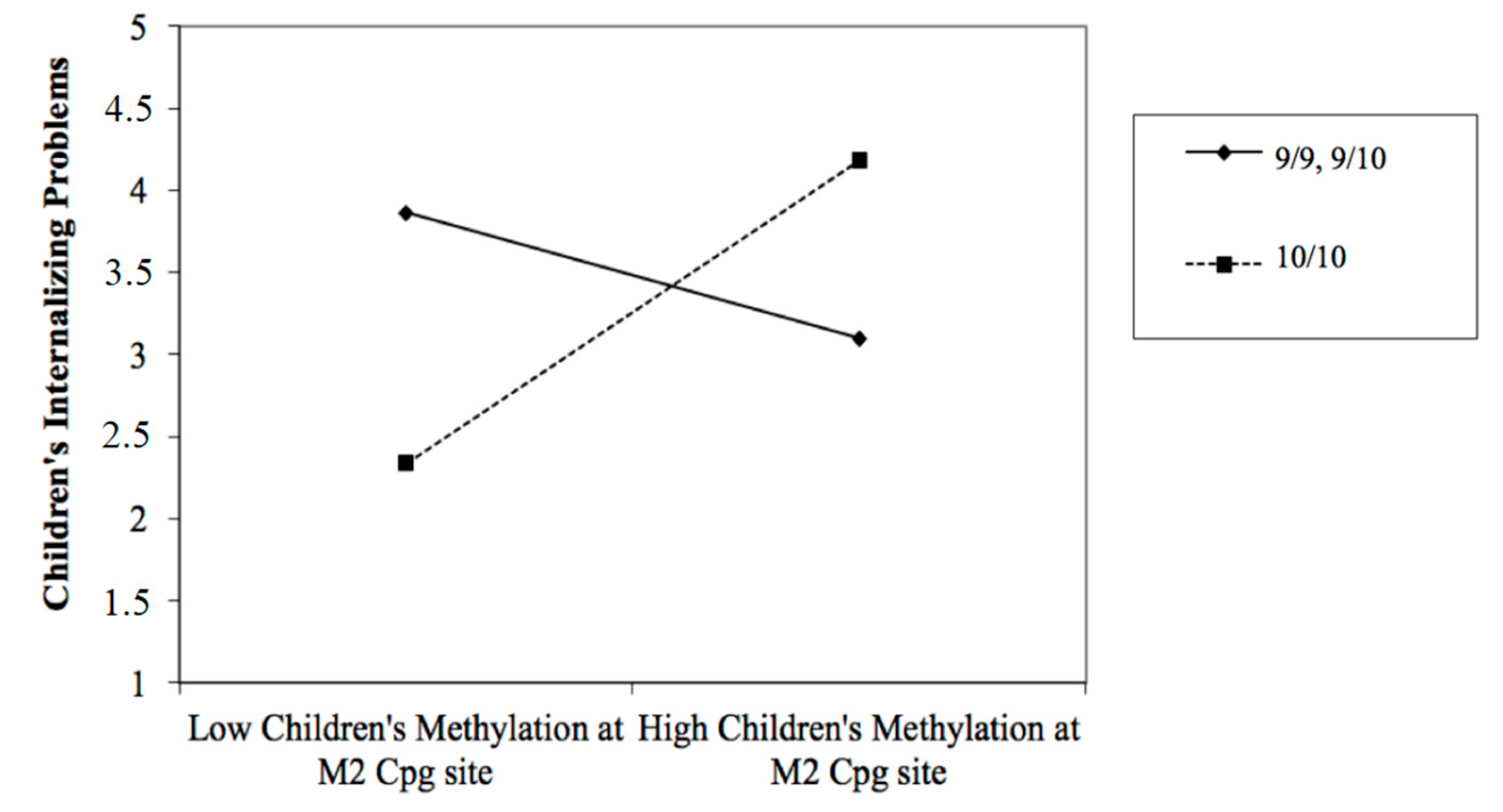
3.2. The Predictive Effect of Children’s Biological Characteristics on Their Own Emotional/Behavioral Functioning, Moderated by DAT1 Genotype
3.3. The Predictive Effect of Parental Psychological Profile and Their Emotional/Behavioral Functioning on Children’s Emotional/Behavioral Functioning, Moderated by Children’s DAT1 Genotype
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- McCrae, R.R.; Costa, P.T., Jr.; Ostendorf, F.; Angleitner, A.; Hřebíčková, M.; Avia, M.D.; Sanz, J.; Sánchez-Bernardos, M.L.; Kusdil, M.E.; Woodfield, R.; et al. Nature over nurture: Temperament, personality, and life span development. J. Personal. Soc. Psychol. 2000, 78, 173. [Google Scholar] [CrossRef]
- Rutter, M.; Moffitt, T.E.; Caspi, A. Gene–environment interplay and psychopathology: Multiple varieties but real effects. J. Child Psychol. Psychiatry 2006, 47, 226–261. [Google Scholar] [CrossRef] [PubMed]
- Sameroff, A. A unified theory of development: A dialectic integration of nature and nurture. Child Dev. 2010, 81, 6–22. [Google Scholar] [CrossRef]
- DiLalla, L.F. Behavior Genetics Principles: Perspectives in Development, Personality, and Psychopathology; American Psychological Association: Washington, DC, USA, 2004. [Google Scholar]
- Oladeji, B.D.; Makanjuola, V.A.; Gureje, O. Family-related adverse childhood experiences as risk factors for psychiatric disorders in Nigeria. Br. J. Psychiatry 2010, 196, 186–191. [Google Scholar] [CrossRef] [PubMed]
- Cimino, S.; Cerniglia, L.; Porreca, A.; Ballarotto, G.; Marzilli, E.; Simonelli, A. Impact of parental binge eating disorder: Exploring children’s emotional/behavioral problems and the quality of parent–child feeding interactions. Infant Ment. Health J. 2018, 39, 552–568. [Google Scholar] [CrossRef] [PubMed]
- Cerniglia, L.; Cimino, S.; Tafà, M.; Marzilli, E.; Ballarotto, G.; Bracaglia, F. Family profiles in eating disorders: Family functioning and psychopathology. Psychol. Res. Behav. Manag. 2017, 10, 305. [Google Scholar] [CrossRef] [PubMed]
- Duncan, L.E.; Keller, M.C. A critical review of the first 10 years of candidate gene-by-environment interaction research in psychiatry. Am. J. Psychiatry 2011, 168, 1041–1049. [Google Scholar] [CrossRef]
- Risch, N.; Herrell, R.; Lehner, T.; Liang, K.Y.; Eaves, L.; Hoh, J.; Griem, A.; Kovacs, M.; Ott, J.; Merikangas, K.R. Interaction between the serotonin transporter gene (5-HTTLPR), stressful life events, and risk of depression: A meta-analysis. JAMA 2009, 301, 2462–2471. [Google Scholar] [CrossRef]
- Belsky, J.; Bakermans-Kranenburg, M.J.; Van IJzendoorn, M.H. For better and for worse: Differential susceptibility to environmental influences. Curr. Dir. Psychol. Sci. 2007, 16, 300–304. [Google Scholar] [CrossRef]
- Belsky, J.; van IJzendoorn, M.H. Genetic differential susceptibility to the effects of parenting. Curr. Opin. Psychol. 2017, 15, 125–130. [Google Scholar] [CrossRef]
- Ellis, B.J.; Boyce, W.T.; Belsky, J.; Bakermans-Kranenburg, M.J.; Van Ijzendoorn, M.H. Differential susceptibility to the environment: An evolutionary–neurodevelopmental theory. Dev. Psychopathol. 2011, 23, 7–28. [Google Scholar] [CrossRef]
- Cicchetti, D.; Rogosch, F.A. Equifinality and multifinality in developmental psychopathology. Dev. Psychopathol. 1996, 8, 597–600. [Google Scholar] [CrossRef] [Green Version]
- Cicchetti, D.; Blender, J.A. A multiple-levels-of-analysis perspective on resilience: Implications for the developing brain, neural plasticity, and preventive interventions. Ann. N. Y. Acad. Sci. 2006, 1094, 248–258. [Google Scholar] [CrossRef]
- Cicchetti, D. Annual research review: Resilient functioning in maltreated children–past, present, and future perspectives. J. Child Psychol. Psychiatry 2013, 54, 402–422. [Google Scholar] [CrossRef]
- Cicchetti, D.; Curtis, W.J. Multilevel perspectives on pathways to resilient functioning. Dev. Psychopathol. 2007, 19, 627–629. [Google Scholar] [CrossRef] [Green Version]
- Masten, A.S.; Cicchetti, D. Developmental cascades. Dev. Psychopathol. 2010, 22, 491–495. [Google Scholar] [CrossRef] [Green Version]
- Rutter, M.; Silberg, J. Gene–environment interplay in relation to emotional and behavioral disturbance. Annu. Rev. Psychol. 2002, 53, 463–490. [Google Scholar] [CrossRef]
- Tambelli, R.; Cerniglia, L.; Cimino, S.; Ballarotto, G. Parent-infant interactions in families with women diagnosed with postnatal depression: A longitudinal study on the effects of a psychodynamic treatment. Front. Psychol. 2015, 6, 1210. [Google Scholar] [CrossRef]
- Cimino, S.; Cerniglia, L.; Ballarotto, G.; Marzilli, E.; Pascale, E.; D’Addario, C.; Adriani, W.; Tambelli, R. DNA Methylation at the DAT Promoter and Risk for Psychopathology: Intergenerational Transmission between School-Age Youths and Their Parents in a Community Sample. Front. Psychiatry 2018, 8, 303. [Google Scholar] [CrossRef] [Green Version]
- Thurston, I.B.; Curley, J.; Fields, S.; Kamboukos, D.; Rojas, A.; Phares, V. How nonclinical are community samples? Am. J. Community Psychol. 2008, 36, 411–420. [Google Scholar] [CrossRef]
- Lavigne, J.V.; Herzing, L.B.; Cook, E.H.; LeBailly, S.A.; Gouze, K.R.; Hopkins, J.; Bryant, F.B. Gene × environment effects of serotonin transporter, dopamine receptor D4, and monoamine oxidase A genes with contextual and parenting risk factors on symptoms of oppositional defiant disorder, anxiety, and depression in a community sample of 4-year-old children. Dev. Psychopathol. 2013, 25, 555–575. [Google Scholar] [CrossRef]
- Nevalainen, N.; Riklund, K.; Andersson, M.; Axelsson, J.; Ögren, M.; Lövdén, M.; Lindenberger, U.; Bäckman, L.; Nyberg, L. COBRA: A prospective multimodal imaging study of dopamine, brain structure and function, and cognition. Brain Res. 2015, 1612, 83–103. [Google Scholar] [CrossRef] [Green Version]
- Vaughan, R.A.; Foster, J.D. Mechanisms of dopamine transporter regulation in normal and disease states. Trends Pharmacol. Sci. 2013, 34, 489–496. [Google Scholar] [CrossRef] [Green Version]
- Friston, K.; Schwartenbeck, P.; FitzGerald, T.; Moutoussis, M.; Behrens, T.; Dolan, R.J. The anatomy of choice: Dopamine and decision-making. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2014, 369, 20130481. [Google Scholar] [CrossRef]
- McHugh, P.C.; Buckley, D.A. The structure and function of the dopamine transporter and its role in CNS diseases. Vitam. Horm. 2015, 8, 339–369. [Google Scholar] [CrossRef]
- Mitchell, R.J.; Howlett, S.; Earl, L.; White, N.G.; McComb, J.; Schanfield, M.S.; Briceno, I.; Papiha, S.S.; Osipova, L.; Livshits, G.; et al. Distribution of the 3’VNTR polymorphism in the human dopamine transporter gene in world populations. Hum. Biol. 2000, 72, 295–304. [Google Scholar] [CrossRef]
- Faraone, S.V.; Bonvicini, C.; Scassellati, C. Biomarkers in the diagnosis of ADHD–promising directions. Curr. Psychiatry Rep. 2014, 16, 497. [Google Scholar] [CrossRef]
- Giana, G.; Romano, E.; Porfirio, M.C.; D’Ambrosio, R.; Giovinazzo, S.; Troianiello, M.; Barlocci, E.; Travaglini, D.; Granstrem, O.; Pascale, E.; et al. Detection of auto-antibodies to DAT in the serum: Interactions with DAT genotype and psycho-stimulant therapy for ADHD. J. Neuroimmunol. 2015, 278, 212–222. [Google Scholar] [CrossRef]
- Pinto-Dussan, M.C.; Avila-Campos, J.E.; Polanco, A.; Vasquez, R.; Arboleda, H. ADHD: Relation between cognitive characteristics and DAT1/DRD4 dopamine polymorphisms. BioRxivorg 2018, 452805. [Google Scholar] [CrossRef]
- Adriani, W.; Pucci, M.; Pascale, E.; Cerniglia, L.; Cimino, S.; Tambelli, R.; Curatolo, P.; Granstrem, O.; Maccarrone, M.; Laviola, G.; et al. Potential for diagnosis versus therapy monitoring of attention deficit hyperactivity disorder: A new epigenetic biomarker interacting with both genotype and auto-immunity. Eur. Child Adolesc. Psychiatry 2018, 27, 241–252. [Google Scholar] [CrossRef]
- Milone, A.; Cerniglia, L.; Cristofani, C.; Inguaggiato, E.; Levantini, V.; Masi, G.; Paciello, M.; Simone, F.; Muratori, P. Empathy in youths with conduct disorder and callous-unemotional traits. Neural Plast 2019, 2019, 9638973. [Google Scholar] [CrossRef]
- Drury, S.S.; Theall, K.P.; Keats, B.J.; Scheeringa, M. The role of the dopamine transporter (DAT) in the development of PTSD in preschool children. J. Trauma. Stress 2009, 22, 534–539. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.S.; Lahey, B.B.; Waldman, I.; Van Hulle, C.A.; Rathouz, P.; Pelham, W.E.; Loney, J.; Cook, E.H. Association of dopamine transporter genotype with disruptive behavior disorders in an eight-year longitudinal study of children and adolescents. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2007, 144, 310–317. [Google Scholar] [CrossRef]
- Gadow, K.D.; Pinsonneault, J.K.; Perlman, G.; Sadee, W. Association of dopamine gene variants, emotion dysregulation and ADHD in autism spectrum disorder. Res. Dev. Disabil. 2014, 35, 1658–1665. [Google Scholar] [CrossRef] [Green Version]
- Mick, E.; Kim, J.W.; Biederman, J.; Wozniak, J.; Wilens, T.; Spencer, T.; Smoller, J.W.; Faraone, S.V. Family based association study of pediatric bipolar disorder and the dopamine transporter gene (SLC6A3). Am. J. Med. Genet. B Neuropsychiatr. Genet. 2008, 147, 1182–1185. [Google Scholar] [CrossRef]
- Cook, E.H., Jr.; Stein, M.A.; Krasowski, M.D.; Cox, N.J.; Olkon, D.M.; Kieffer, J.E.; Leventhal, B.L. Association of attention-deficit disorder and the dopamine transporter gene. Am. J. Hum. Genet. 1995, 56, 993. [Google Scholar] [CrossRef]
- Gizer, I.R.; Ficks, C.; Waldman, I.D. Candidate gene studies of ADHD: A meta-analytic review. J. Hum. Genet. 2009, 126, 51–90. [Google Scholar] [CrossRef]
- Anreiter, I.; Sokolowski, H.M.; Sokolowski, M.B. Gene–environment interplay and individual differences in behavior. Mind Brain Educ. 2018, 12, 200–211. [Google Scholar] [CrossRef]
- Roth, T.L. Epigenetic mechanisms in the development of behavior: Advances, challenges, and future promises of a new field. Dev. Psychopathol. 2013, 25, 1279–1291. [Google Scholar] [CrossRef] [Green Version]
- Tronick, E.; Hunter, R.G. Waddington, dynamic systems, and epigenetics. Front. Behav. Neurosci. 2016, 10, 107. [Google Scholar] [CrossRef]
- Shanahan, M.J.; Hofer, S.M. Social context in gene–environment interactions: Retrospect and prospect. J. Gerontol. B Psychol. Sci. Soc. Sci. 2005, 60, 65–76. [Google Scholar] [CrossRef]
- Sonuga-Barke, E.J.; Oades, R.D.; Psychogiou, L.; Chen, W.; Franke, B.; Buitelaar, J.; Banaschewski, T.; Ebstein, R.P.; Gil, M.; Anney, R.; et al. Dopamine and serotonin transporter genotypes moderate sensitivity to maternal expressed emotion: The case of conduct and emotional problems in attention deficit/hyperactivity disorder. J. Child Psychol. Psychiatry 2009, 50, 1052–1063. [Google Scholar] [CrossRef]
- Li, J.J.; Lee, S.S. Interaction of dopamine transporter gene and observed parenting behaviors on attention-deficit/hyperactivity disorder: A structural equation modeling approach. J. Clin. Child Adolesc. Psychol. 2013, 42, 174–186. [Google Scholar] [CrossRef]
- Hayden, E.P.; Hanna, B.; Sheikh, H.I.; Laptook, R.S.; Kim, J.; Singh, S.M.; Klein, D.N. Child dopamine active transporter 1 genotype and parenting: Evidence for evocative gene–environment correlations. Dev. Psychopathol. 2013, 25, 163–173. [Google Scholar] [CrossRef]
- Villani, V.; Ludmer, J.; Gonzalez, A.; Levitan, R.; Kennedy, J.; Masellis, M.; Basile, V.S.; Wekerle, C.; Atkinson, L. Dopamine receptor D2 (DRD2), dopamine transporter solute carrier family C6, member 4 (SLC6A3), and catechol-O-methyltransferase (COMT) genes as moderators of the relation between maternal history of maltreatment and infant emotion regulation. Dev. Psychopathol. 2018, 30, 581–592. [Google Scholar] [CrossRef]
- Caspi, A.; McClay, J.; Moffitt, T.E.; Mill, J.; Martin, J.; Craig, I.W.; Taylor, A.; Poulton, R. Role of genotype in the cycle of violence in maltreated children. Science 2002, 297, 851–854. [Google Scholar] [CrossRef]
- Champagne, F.A.; Curley, J.P. Epigenetic mechanisms mediating the long-term effects of maternal care on development. Neurosci. Biobehav. Rev. 2009, 33, 593–600. [Google Scholar] [CrossRef]
- Schuch, V.; Utsumi, D.A.; Costa, T.V.M.M.; Kulikowski, L.D.; Muszkat, M. Attention deficit hyperactivity disorder in the light of the epigenetic paradigm. Front. Psychiatry 2015, 6, 126. [Google Scholar] [CrossRef]
- Perroud, N.; Zewdie, S.; Stenz, L.; Adouan, W.; Bavamian, S.; Prada, P.; Nicastro, R.; Hasler, R.; Nallet, A.; Piguet, C.; et al. Methylation of serotonin receptor 3A in ADHD, borderline personality, and bipolar disorders: Link with severity of the disorders and childhood maltreatment. Depress. Anxiety 2016, 33, 45–55. [Google Scholar] [CrossRef]
- Lopizzo, N.; Bocchio Chiavetto, L.; Cattane, N.; Plazzotta, G.; Tarazi, F.I.; Pariante, C.M.; Riva, M.A.; Cattaneo, A. Gene–environment interaction in major depression: Focus on experience-dependent biological systems. Front. Psychiatry 2015, 6, 68. [Google Scholar] [CrossRef]
- McKay, J.A.; Groom, A.; Potter, C.; Coneyworth, L.J.; Ford, D.; Mathers, J.C.; Relton, C.L. Genetic and non-genetic influences during pregnancy on infant global and site specific DNA methylation: Role for folate gene variants and vitamin B12. PLoS ONE 2012, 7, e33290. [Google Scholar] [CrossRef] [PubMed]
- Lövkvist, C.; Dodd, I.B.; Sneppen, K.; Haerter, J.O. DNA methylation in human epigenomes depends on local topology of CpG sites. Nucleic Acids Res. 2016, 44, 5123–5132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Lior-Hoffmann, L.; Wang, S.; Zhang, Y.; Broyde, S. DNA cytosine methylation: Structural and thermodynamic characterization of the epigenetic marking mechanism. Biochemistry 2013, 52, 2828–2838. [Google Scholar] [CrossRef] [PubMed]
- Shumay, E.; Fowler, J.S.; Volkow, N.D. Genomic features of the human dopamine transporter gene and its potential epigenetic States: Implications for phenotypic diversity. PLoS ONE 2010, 5, e11067. [Google Scholar] [CrossRef] [PubMed]
- Jasiewicz, A.; Rubiś, B.; Samochowiec, J.; Małecka, I.; Suchanecka, A.; Jabłoński, M.; Grzywacz, A. DAT1 methylation changes in alcohol-dependent individuals vs. controls. J. Psychiatr. Res. 2015, 64, 130–133. [Google Scholar] [CrossRef] [PubMed]
- Wiers, C.E.; Shumay, E.; Volkow, N.D.; Frieling, H.; Kotsiari, A.; Lindenmeyer, J.; Walter, H.; Bermpohl, F. Effects of depressive symptoms and peripheral DAT methylation on neural reactivity to alcohol cues in alcoholism. Transl. Psychiatry 2015, 5, e648. [Google Scholar] [CrossRef]
- Ding, K.; Yang, J.; Reynolds, G.P.; Chen, B.; Shao, J.; Liu, R.; Yang, R.; Wen, J.; Kang, C. DAT1 methylation is associated with methylphenidate response on oppositional and hyperactive-impulsive symptoms in children and adolescents with ADHD. World J. Biol. Psychiatry 2017, 18, 291–299. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Chen, X.T.; Luo, M.; Tang, Y.; Zhang, G.; Wu, D.; Yang, B.; Ruan, D.Y.; Wang, H.L. Multiple epigenetic factors predict the attention deficit/hyperactivity disorder among the Chinese Han children. J. Psychiatr. Res. 2015, 64, 40–50. [Google Scholar] [CrossRef] [PubMed]
- Giedd, J.N. The teen brain: Insights from neuroimaging. J. Adolesc. Health 2008, 42, 335–343. [Google Scholar] [CrossRef]
- Mills, K.L.; Lalonde, F.; Clasen, L.S.; Giedd, J.N.; Blakemore, S.J. Developmental changes in the structure of the social brain in late childhood and adolescence. Soc. Cogn. Affect. Neurosci. 2012, 9, 123–131. [Google Scholar] [CrossRef]
- Lucarelli, M.; Visalberghi, E.; Adriani, W.; Addessi, E.; Pierandrei, S.; Manciocco, A.; Zoratto, F.; Tamellini, A.; Vitale, A.; Laviola, G.; et al. Polymorphism of the 3’-UTR of the dopamine transporter gene (DAT) in New World monkeys. Primates 2017, 58, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Lahey, B.B.; Rathouz, P.J.; Lee, S.S.; Chronis-Tuscano, A.; Pelham, W.E.; Waldman, I.D.; Cook, E.H. Interactions between early parenting and a polymorphism of the child’s dopamine transporter gene in predicting future child conduct disorder symptoms. J. Abnorm. Psychol. 2011, 120, 33. [Google Scholar] [CrossRef] [PubMed]
- Palma-Gudiel, H.; Fañanás, L. An integrative review of methylation at the serotonin transporter gene and its dialogue with environmental risk factors, psychopathology and 5-HTTLPR. Neurosci. Biobehav. Rev. 2017, 72, 190–209. [Google Scholar] [CrossRef] [PubMed]
- Faraone, S.V.; Mick, E. Molecular genetics of attention deficit hyperactivity disorder. Psychiatr. Clin. N. Am. 2010, 33, 159–180. [Google Scholar] [CrossRef] [PubMed]
- van de Giessen, E.M.; de Win, M.M.; Tanck, M.W.; van den Brink, W.; Baas, F.; Booij, J. Striatal dopamine transporter availability associated with polymorphisms in the dopamine transporter gene SLC6A3. J. Nucl. Med. 2009, 50, 45–52. [Google Scholar] [CrossRef]
- Fitzmaurice, G.M.; Laird, N.M.; Ware, J.H. Applied Longitudinal Analysis; John Wiley & Sons: Hoboken, NJ, USA, 2012. [Google Scholar]
- Cox, D.R. Causality: Some Statistical Aspects. J. R. Stat. Soc. Ser. A Stat. Soc. 1992, 155, 291–301. [Google Scholar] [CrossRef]
- Wunsch, G.; Russo, F.; Mouchart, M. Do we necessarily need longitudinal data to infer causal relations? Bull. Sociol. Methodol. 2010, 106, 5–18. [Google Scholar] [CrossRef]
- Reichenheim, M.E.; Coutinho, E.S. Measures and models for causal inference in cross-sectional studies: Arguments for the appropriateness of the prevalence odds ratio and related logistic regression. BMC Med. Res. Methodol. 2010, 10, 66. [Google Scholar] [CrossRef]
- Hollingshead, A.B. Four Factor Index of Social Status; Yale University: New Haven, CT, USA, 1975. [Google Scholar]
- Torrone, D.Z.; Kuriakose, J.S.; Moors, K.; Jiang, H.; Niedzwiecki, M.M.; Perera, F.F.; Miller, R.L. Reproducibility and intraindividual variation over days in buccal cell DNA methylation of two asthma genes, interferon γ (IFNγ) and inducible nitric oxide synthase (iNOS). Clin. Epigenet. 2012, 4, 3. [Google Scholar] [CrossRef]
- Prunas, A.; Sarno, I.; Preti, E.; Madeddu, F.; Perugini, M. Psychometric properties of the Italian version of the SCL-90-R: A study on a large community sample. Eur. Psychiatry 2012, 27, 591–597. [Google Scholar] [CrossRef]
- Cimino, S.; Cerniglia, L.; Dentale, F.; Capobianco, M.; Tambelli, R. Maternal symptoms of depression and paranoid ideation can be predictive of the onset of eating disorders in early adolescents offsprings: A nine-year longitudinal study. Int. J. Psychol. Psychol. Ther. 2018, 18, 221–234. [Google Scholar]
- Deragotis, L.R. Symptom Checklist-90-R: Administration, Scoring, and Procedures Manual; National Computer Systems: Minneapolis, MN, USA, 1994. [Google Scholar]
- Achenbach, T.M.; Rescorla, L.A. Manual for the ASEBA Adult Forms & Profiles; Research Center for Children, Youth, & Families: Burlington, VT, USA, 2003. [Google Scholar]
- Achenbach, T.M.; Rescorla, L.A. Manual for the ASEBA School-Age Forms & Profiles; Research Center for Children, Youth, & Families: Burlington, VT, USA, 2001. [Google Scholar]
- Frigerio, A.; Cattaneo, C.; Cataldo, M.; Schiatti, A.; Molteni, M.; Battaglia, M. Behavioral and emotional problems among Italian children and adolescents aged 4 to 18 years as reported by parents and teachers. Eur. J. Psychol. Assess. 2004, 20, 124–133. [Google Scholar] [CrossRef]
- Hay, D.F.; Pawlby, S.; Sharp, D.; Schmücker, G.; Mills, A.; Allen, H.; Kumar, R. Parents’ judgements about young children’s problems: Why mothers and fathers might disagree yet still predict later outcomes. J. Child Psychol. Psychiatry 1999, 40, 1249–1258. [Google Scholar] [CrossRef] [PubMed]
- Christensen, A.; Margolin, G.; Sullaway, M. Interparental agreement on child behavior problems. Psychol. Assess. 1992, 4, 419. [Google Scholar] [CrossRef]
- Jansen, M.; Bodden, D.H.; Muris, P.; van Doorn, M.; Granic, I. Measuring anxiety in children: The importance of separate mother and father reports. Child Youth Care Forum 2017, 46, 643–659. [Google Scholar] [CrossRef] [PubMed]
- Weijers, D.; van Steensel, F.J.A.; Bögels, S.M. Associations between psychopathology in mothers, fathers and their children: A structural modeling approach. J. Child Fam. Stud. 2018, 27, 1992–2003. [Google Scholar] [CrossRef] [PubMed]
- Purcell, S. Variance components models for gene–environment interaction in twin analysis. Twin Res. Hum. Genet. 2002, 5, 554–571. [Google Scholar] [CrossRef] [PubMed]
- Wermter, A.K.; Laucht, M.; Schimmelmann, B.G.; Banaschweski, T.; Sonuga-Barke, E.J.; Rietschel, M.; Becker, K. From nature versus nurture, via nature and nurture, to gene× environment interaction in mental disorders. Eur. Child Adolesc. Psychiatry 2010, 19, 199–210. [Google Scholar] [CrossRef] [PubMed]
- Richards, J.S.; Arias Vásquez, A.; van Rooij, D.; van der Meer, D.; Franke, B.; Hoekstra, P.J.; Heslenfeld, D.J.; Oosterlaan, J.; Faraone, S.V.; Hartman, C.A.; et al. Testing differential susceptibility: Plasticity genes, the social environment, and their interplay in adolescent response inhibition. World J. Biol. Psychiatry 2017, 18, 308–321. [Google Scholar] [CrossRef] [PubMed]
- Hayes, A.F. Introduction to Mediation, Moderation, and Conditional Process Analysis: A Regression-Based Approach; Guilford Publications: New York, NY, USA, 2017. [Google Scholar]
- Hayes, A.F.; Scharkow, M. The relative trustworthiness of inferential tests of indirect effect in statistical mediation analysis: Does method really matters? Psychol. Sci. 2013, 24, 1918–1927. [Google Scholar] [CrossRef] [PubMed]
- Aiken, L.S.; West, S.G.; Reno, R.R. Multiple Regression: Testing and Interpreting Interactions; Sage: Thousand Oaks, CA, USA, 1991. [Google Scholar]
- Rathouz, P.J.; Van Hulle, C.A.; Rodgers, J.L.; Waldman, I.; Lahey, B.B. Specification, testing, and interpretation of gene-by-measured-environment interaction models in the presence of gene-environment correlation. Behav. Genet. 2008, 38, 301–315. [Google Scholar] [CrossRef] [PubMed]
- Beach, S.R.; Brody, G.H.; Barton, A.W.; Philibert, R.A. Exploring genetic moderators and epigenetic mediators of contextual and family effects: From Gene× Environment to epigenetics. Dev. Psychopathol. 2016, 28, 1333–1346. [Google Scholar] [CrossRef] [PubMed]
- Belsky, J. The differential susceptibility hypothesis: Sensitivity to the environment for better and for worse. JAMA Pediatr. 2016, 170, 321–322. [Google Scholar] [CrossRef] [PubMed]
- Barker, E.D.; Walton, E.; Cecil, C.A. Annual Research Review: DNA methylation as a mediator in the association between risk exposure and child and adolescent psychopathology. J. Child Psychol. Psychiatry 2018, 59, 303–322. [Google Scholar] [CrossRef] [PubMed]
- Gaunt, T.R.; Shihab, H.A.; Hemani, G.; Min, J.L.; Woodward, G.; Lyttleton, O.; Zheng, J.; Duggirala, A.; McArdle, W.L.; Ho, K.; et al. Systematic identification of genetic influences on methylation across the human life course. Genome Biol. 2016, 17, 61. [Google Scholar] [CrossRef] [PubMed]
- Teh, A.L.; Pan, H.; Chen, L.; Ong, M.L.; Dogra, S.; Wong, J.; MacIsaac, J.L.; Mah, S.M.; McEwen, L.M.; Saw, S.M.; et al. The effect of genotype and in utero environment on interindividual variation in neonate DNA methylomes. Genome Res. 2014, 24, 1064–1074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cimino, S.; Cerniglia, L.; Porreca, A.; Simonelli, A.; Ronconi, L.; Ballarotto, G. Mothers and fathers with binge eating disorder and their 18–36 months old children: A longitudinal study on parent–infant interactions and offspring’s emotional–behavioral profiles. Front. Psychol. 2016, 7, 580. [Google Scholar] [CrossRef] [PubMed]
- Tafà, M.; Cimino, S.; Ballarotto, G.; Bracaglia, F.; Bottone, C.; Cerniglia, L. Female adolescents with eating disorders, parental psychopathological risk and family functioning. J. Child Fam. Stud. 2017, 26, 28–39. [Google Scholar] [CrossRef]
- Hughes, V. Epigenetics: The sins of the father. Nature 2014, 507, 22. [Google Scholar] [CrossRef]
- Champagne, F.A.; Mashoodh, R. Genes in context: Gene–environment interplay and the origins of individual differences in behavior. Curr. Dir. Psychol. Sci. 2009, 18, 127–131. [Google Scholar] [CrossRef]
- Burt, S.A. Some key issues in the study of gene-environment interplay: Activation, deactivation, and the role of development. Res. Hum. Dev. 2011, 8, 192–210. [Google Scholar] [CrossRef]
- van IJzendoorn, M.H.; Caspers, K.; Bakermans-Kranenburg, M.J.; Beach, S.R.; Philibert, R. Methylation matters: Interaction between methylation density and serotonin transporter genotype predicts unresolved loss or trauma. Biol. Psychiatry 2010, 68, 405–407. [Google Scholar] [CrossRef] [PubMed]
- Parade, S.H.; Ridout, K.K.; Seifer, R.; Armstrong, D.A.; Marsit, C.J.; McWilliams, M.A.; Tyrka, A.R. Methylation of the glucocorticoid receptor gene promoter in preschoolers: Links with internalizing behavior problems. Child Dev. 2016, 87, 86–97. [Google Scholar] [CrossRef] [PubMed]
- Cimino, S.; Cerniglia, L.; Paciello, M. Mothers with depression, anxiety or eating disorders: Outcomes on their children and the role of paternal psychological profiles. Child Psychiatry Hum. D 2015, 46, 228–236. [Google Scholar] [CrossRef]
- Middeldorp, C.M.; Wesseldijk, L.W.; Hudziak, J.J.; Verhulst, F.C.; Lindauer, R.J.; Dieleman, G.C. Parents of children with psychopathology: Psychiatric problems and the association with their child’s problems. Eur. Child Adolesc. Psychiatry 2016, 25, 919–927. [Google Scholar] [CrossRef]
- Pluess, M.; Velders, F.P.; Belsky, J.; van IJzendoorn, M.H.; Bakermans-Kranenburg, M.J.; Jaddoe, V.W.; Hofman, A.; Arp, P.P.; Verhulst, F.C.; Tiemeier, H. Serotonin transporter polymorphism moderates effects of prenatal maternal anxiety on infant negative emotionality. Biol. Psychiatry 2011, 69, 520–525. [Google Scholar] [CrossRef] [PubMed]
- Gibb, B.E.; Uhrlass, D.J.; Grassia, M.; Benas, J.S.; McGeary, J. Children’s inferential styles, 5-HTTLPR genotype, and maternal expressed emotion-criticism: An integrated model for the intergenerational transmission of depression. J. Abnorm. Psychol. 2009, 118, 734. [Google Scholar] [CrossRef] [PubMed]
- Stevens, S.E. Adolescent Inattention/Overactivity/Impulsivity as an Outcome of Early Institutional Deprivation: The Role of Genetic Factors. Ph.D. Thesis, University of Southampton, Southampton, UK, 2009. [Google Scholar]
- Papp, L.M.; Goeke-Morey, M.C.; Cummings, E.M. Mothers’ and fathers’ psychological symptoms and marital functioning: Examination of direct and interactive links with child adjustment. J. Child Fam. Stud. 2004, 13, 469–482. [Google Scholar] [CrossRef]
- Camisasca, E.; Miragoli, S.; Di Blasio, P. Families with distinct levels of marital conflict and child adjustment: Which role for maternal and paternal stress? J. Child Fam. Stud. 2016, 25, 733–745. [Google Scholar] [CrossRef]
- Schleider, J.L.; Ginsburg, G.S.; Keeton, C.P.; Weisz, J.R.; Birmaher, B.; Kendall, P.C.; Piacentini, J.; Sherrill, J.; Walkup, J.T. Parental psychopathology and treatment outcome for anxious youth: Roles of family functioning and caregiver strain. J. Consult. Clin. Psychol. 2015, 83, 213. [Google Scholar] [CrossRef]
- Bellgrove, M.A.; Chambers, C.D.; Johnson, K.A.; Daibhis, A.; Daly, M.; Hawi, Z.; Lambert, D.; Gill, M.; Robertson, I.H. Dopaminergic genotype biases spatial attention in healthy children. Mol. Psychiatry 2007, 12, 786. [Google Scholar] [CrossRef] [PubMed]
- Boyce, W.T.; Kobor, M.S. Development and the epigenome: The ‘synapse’of gene–environment interplay. Dev. Sci. 2015, 18, 1–23. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Predictors | Outcome: CBCL/6-18_Internalizing Problems | |||||
|---|---|---|---|---|---|---|
| Model 1 | Model 2 | |||||
| B | t | p | B | t | p | |
| M1 | −0.35 | −1.61 | 0.11 | −0.15 | −0.44 | 0.66 |
| M2 | 1.33 | 3.25 | 0.00 ** | 0.05 | 0.07 | 0.94 |
| M3 | −0.59 | −1.60 | 0.11 | 0.44 | 0.58 | 0.56 |
| M5 | −0.51 | −1.69 | 0.10 | −0.23 | −0.45 | 0.65 |
| M6 | 0.60 | 2.16 | 0.03 * | 0.55 | 1.39 | 0.17 |
| M7 | −0.12 | −0.59 | 0.55 | −0.33 | −1.09 | 0.28 |
| DAT1 genotype a | 0.08 | 0.55 | 0.58 | −0.23 | −0.96 | 0.34 |
| DAT1 × M1 | −0.04 | −0.16 | 0.86 | |||
| DAT1 × M2 | 1.76 | 2.22 | 0.03 * | |||
| DAT1 × M3 | −1.33 | −1.75 | 0.09 | |||
| DAT1 × M5 | −0.21 | −0.4 | 0.69 | |||
| DAT1 × M6 | 0.68 | 0.94 | 0.35 | |||
| DAT1 × M7 | −0.39 | −0.84 | 0.40 | |||
| R2 | 0.495 | 0.597 | ||||
| Predictors | Outcome: CBCL/6-18_ Externalizing Problems | |||||
|---|---|---|---|---|---|---|
| Model 1 | Model 2 | |||||
| B | t | p | B | t | p | |
| ASR Internalizing mothers | 0.15 | 0.90 | 0.37 | 0.13 | 0.64 | 0.52 |
| ASR Externalizing mothers | 0.20 | 1.61 | 0.11 | 0.18 | 1,2 | 0.22 |
| SCL-90-R GSI mothers | −0.02 | −0.16 | 0.87 | 0.12 | 0.57 | 0.56 |
| ASR Internalizing fathers | −0.29 | −1.35 | 0.17 | −0.47 | −1.42 | 0.07 |
| ASR Externalizing fathers | 0.16 | 0.89 | 0.37 | 0.51 | 2.20 | 0.03 * |
| SCL-90-R GSI fathers | 0.23 | 1.18 | 0.24 | 0.20 | 0.89 | 0.37 |
| DAT1 genotype a | −0.14 | −1.22 | 0.22 | −0.11 | −0.97 | 0.33 |
| DAT1 × Internalizing mothers | 0.01 | 0.07 | 0.94 | |||
| DAT1 × Externalizing mothers | −0.01 | −0.07 | 0.94 | |||
| DAT1 × GSI mothers | −0.16 | −0.70 | 0.48 | |||
| DAT1 × Internalizing fathers | 0.30 | 1.32 | 0.19 | |||
| DAT1 × Externalizing fathers | −0.60 | −2.07 | 0.04 * | |||
| DAT1 × GSI fathers | 0.09 | 0.44 | 0.66 | |||
| R2 | 0.145 | 0.23 | ||||
| Predictors | Outcome: CBCL/6-18_ Total Problems | |||||
|---|---|---|---|---|---|---|
| Model 1 | Model 2 | |||||
| B | t | p | B | t | p | |
| ASR Internalizing mothers | 0.01 | 0.09 | 0.92 | 0.04 | 0.22 | 0.82 |
| ASR Externalizing mothers | 0.11 | 0.94 | 0.34 | 0.20 | 1.34 | 0.18 |
| SCL-90-R GSI mothers | 0.30 | 1.93 | 0.05 | 0.21 | 1.01 | 0.31 |
| ASR Internalizing fathers | −0.13 | −0.64 | 0.52 | −0.25 | −1.01 | 0.31 |
| ASR Externalizing fathers | 0.21 | 1.21 | 0.23 | −0.45 | 2.02 | 0.04 * |
| SCL-90-R GSI fathers | 0.11 | 0.60 | 0.55 | 0.08 | 0.36 | 0.71 |
| DAT1 genotype a | −0.19 | −1.72 | 0.09 | −0.17 | −1.53 | 0.13 |
| DAT1 × Internalizing mothers | −0.05 | −0.22 | 0.82 | |||
| DAT1 × Externalizing mothers | −0.20 | −1.41 | 0.16 | |||
| DAT1 × GSI mothers | 0.23 | 1.01 | 0.31 | |||
| DAT1 × Internalizing fathers | 0.10 | 0.48 | 0.62 | |||
| DAT1 × Externalizing fathers | −0.48 | −1.71 | 0.09 | |||
| DAT1 × GSI fathers | 0.15 | 0.73 | 0.46 | |||
| R2 | 0.224 | 0.29 | ||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cimino, S.; Cerniglia, L.; Ballarotto, G.; Marzilli, E.; Pascale, E.; D’Addario, C.; Adriani, W.; Maremmani, A.G.I.; Tambelli, R. Children’s DAT1 Polymorphism Moderates the Relationship Between Parents’ Psychological Profiles, Children’s DAT Methylation, and Their Emotional/Behavioral Functioning in a Normative Sample. Int. J. Environ. Res. Public Health 2019, 16, 2567. https://doi.org/10.3390/ijerph16142567
Cimino S, Cerniglia L, Ballarotto G, Marzilli E, Pascale E, D’Addario C, Adriani W, Maremmani AGI, Tambelli R. Children’s DAT1 Polymorphism Moderates the Relationship Between Parents’ Psychological Profiles, Children’s DAT Methylation, and Their Emotional/Behavioral Functioning in a Normative Sample. International Journal of Environmental Research and Public Health. 2019; 16(14):2567. https://doi.org/10.3390/ijerph16142567
Chicago/Turabian StyleCimino, Silvia, Luca Cerniglia, Giulia Ballarotto, Eleonora Marzilli, Esterina Pascale, Claudio D’Addario, Walter Adriani, Angelo Giovanni Icro Maremmani, and Renata Tambelli. 2019. "Children’s DAT1 Polymorphism Moderates the Relationship Between Parents’ Psychological Profiles, Children’s DAT Methylation, and Their Emotional/Behavioral Functioning in a Normative Sample" International Journal of Environmental Research and Public Health 16, no. 14: 2567. https://doi.org/10.3390/ijerph16142567