

Abstract
Background: Suicidal behaviour aggregates in families, and the hypothalamic–pituitary–adrenal (HPA) axis and noradrenergic dysregulation may play a role in suicide risk. It is unclear whether stress dysregulation is a heritable trait of suicide or how it might increase risk. We investigated stress reactivity of the autonomic nervous system and the HPA axis in suicide predisposition and characterized the effect of this dysregulation on neuropsychologic function.
Methods: In this family-based study of first-degree relatives (n = 14) of suicide completers and matched controls with no family or personal history of suicidal behaviour (n = 14), participants underwent the Trier Social Stress Test (TSST). We used salivary α-amylase and cortisol levels to characterize stress reactivity and diurnal variation. We administered a series of neuropsychologic and executive function tests before and after the TSST.
Results: Despite normal diurnal variation, relatives of suicide completers exhibited blunted cortisol and α-amylase TSST reactivity. Although there were no baseline differences in conceptual reasoning, sustained attention or executive function, the relatives of suicide completers did not improve on measures of inhibition upon repeated testing after TSST. Secondary analyses suggested that these effects were related to suicide vulnerability independent of major depression.
Limitations: The sample size was small, and the design prevents us from disentangling our findings from the possible traumatic consequences of losing a relative by suicide.
Conclusions: Blunted stress response may be a trait of suicide risk, and impairment of stress-induced executive function may contribute to suicide vulnerability.
Introduction
Suicide is one of the leading causes of death in Western countries.1 Well-investigated suicide risk factors include suicide attempts, psychopathology, certain demographic variables and psychosocial factors, such as negative life events and a lack of social support.2,3
It is well established that suicide aggregates in families.2,4–7 Although several studies have suggested the involvement of familial and heritable personality factors, such as impulsive aggression,4,7,8 our understanding of the mechanisms of transmission within families is limited.
Suicide is likely the result of interactions among biological factors; these interactions increase an individual’s predisposition to suicidal behaviour, psychopathology and environmental stressors. Neurocognitive function and the physiologic response to stress are 2 promising intermediate phenotypes that may underlie the familial transmission of suicide.9 Investigations using family-based study designs are therefore important to test their role and potential interaction in suicide vulnerability.
Dysregulation of the hypothalamic–pituitary–adrenal (HPA) axis is hypothesized to play a role in increasing susceptibility to suicidal behaviour.10 The role of dysregulation of the HPA axis as an individual susceptibility factor for suicide has been supported by pharmacologic studies using the dexamethasone suppression test,11–17 with several studies indicating that dexamethasone nonsuppression is related to increased risk of suicidal behaviour. The relation of glucocorticoid secretion to suicide is clinically relevant, with potential implications in the etiology of suicide through its well-characterized association with stress. However, the dynamic changes in indicators of biological reactivity to stress in relation to suicide have not been well characterized, and the subjective experience of stress has not been not well modelled using pharmacologic manipulation of the stress systems.
The Trier Social Stress Test (TSST18) is a well-investigated psychosocial stress paradigm that activates the HPA axis and the sympathetic nervous system.19,20 The TSST allows repeated salivary assessments throughout the test to quantify the dynamic changes in biological markers of stress. Given the consistent evidence suggesting that a portion of the variability in TSST reactivity is because of genetic factors,21,22 this method is well suited to an investigation of first-degree relatives to determine the role of stress reactivity as an intermediate phenotype of suicide.
To further explore our hypothesis that stress dysregulation is a heritable risk factor for suicidal behaviour, we conducted neuropsychologic testing focusing on executive function to determine whether this is a psychological process in which this increased vulnerability is manifested. Executive function is an important superordinate cognitive ability. With deficits classically associated with frontal lobe damage, executive function is involved in important domains such as cognitive flexibility as well as initiating appropriate actions and inhibiting inappropriate actions. In patients with major depressive disorder, neuropsychologic testing has revealed the importance of deficits in executive function in suicidal behaviour with the Wisconsin Card Sorting Test,23 Iowa Gambling Task24 and go/no-go inhibition tasks.25 The inability to adapt to changing environments, see alternatives and inhibit inappropriate or maladaptive behaviours is of important clinical relevance for suicide.
Situational stressors are associated with a range of consequences for cognitive performance.26 With the growing literature in humans using exogenous corticosteroids27 and noradrenergic function28,29 in modulating executive function, we sought to investigate the impact of a controlled laboratory psychosocial stress on executive function in a vulnerable population.
To investigate stress reactivity and neurocognitive function as mechanisms of familial transmission of suicide, we included first-degree relatives of people who had committed suicide (hereinafter called relatives) and a matched population controls without a family or personal history of suicidal behaviour. We hypothesized that the relatives would show significant differences in the response of the HPA axis and sympathetic nervous system to social stress, without affected diurnal variation. We also characterized the neuropsychologic consequences of psychosocial stress for their role in suicide risk and hypothesized a role between stress and suicide risk.
Methods
Study population
Through collaboration with the Quebec Coroner’s Office, we were able to characterize a representative sample of people who had committed suicide in the Montréal region. We included first-degree relatives of people who had committed suicide (n = 14). None of the relatives currently had relevant psychopathology, including depressive disorders, substance disorders or posttraumatic stress disorder. Relatives did not work night shifts nor were they involved in a profession that required frequent public speaking (e.g., politicians, lawyers).
We identified healthy controls (n = 14) through advertisements in local newspapers. Controls were included after semistructured interviews confirmed the absence of a personal or a family history of suicidal behaviour and relevant psychopathology, including depressive disorders, substance disorders and posttraumatic stress disorder. Controls did not work night shifts nor were they involved in a profession requiring frequent public speaking.
This study was approved by the research ethics board of the Douglas University Mental Health Institute. Relatives and controls signed informed consent forms.
Clinical assessment
All participants underwent psychiatric assessment before neuropsychologic and stress testing. Psychiatric disorders were assessed by use of the Structured Clinical Interview for DSM-IV Axis I30 and Axis II31 disorders. We deemed participants eligible if these schedules revealed the absence of current psychopathology. This was important given the important alterations in neuroendocrine and neurocognitive function associated with affective disorders, substance use disorders and posttraumatic stress disorder.
All participants were instructed not to consume alcohol the day before or on the day of testing. Testing took place between 1 and 5 pm, when participants also completed the Beck Depression Inventory (BDI)32 and Beck Anxiety Inventory (BAI) between 1 and 5 pm.33
Trier Social Stress Test
Participants underwent psychosocial stress testing with a modified version of the TSST.18 Participants were informed that they would give a 10-minute improvised speech about personal strengths and weaknesses; they were given 10 minutes to prepare a speech. They were informed that they would be observed by a psychiatrist (G.D. or M.T.B.) trained in behavioural analysis. Unbeknownst to the participants, the true role of the observer was to maintain a neutral facial expression, give neither positive nor negative feedback at any time during the presentation and take notes to increase participants’ self-awareness. The observer was unaware of whether the participants were relatives or controls. Five minutes into the presentation, participants were instructed by the observer to perform serial subtractions of 17 starting at 2041; participants were asked to start again if they made a mistake and were pressed to continue if they paused for 10 seconds.
To test for variation in salivary cortisol and α-amylase levels, we collected 8 saliva samples per participant using salivettes (Sarstedt) every 10 minutes starting 40 minutes before the test (−40, −30, −20, −10 min/anticipation [ANT]) and 30 minutes after the TSST.
Neuropsychologic testing
We administered the following, well-investigated neuropsychologic tests at baseline (30 min before the saliva samples were collected): the Wechsler Adult Intelligence Scale (WAIS) III, Matrix Reasoning,34 the Continuous Performance Test (CPT35) and the Delis–Kaplan Executive Function System (D-KEFS36).
The WAIS includes a test of abstract reasoning that presents a series of geometrical shapes; participants must select 1 of 5 possibilities that respect the abstract rule. The CPT is a computerized test that requires sustained attention to 450 numbers composed of 4 digits that are briefly flashed on the screen. Participants monitor for identical repeated numbers. This test provides correct responses and false alarms. The D-KEFS is a series of tests focused on different aspects of executive function. It provides scaled scores, with higher scores representing better performance (i.e., higher completion time scores indicate shorter completion times, and higher error scores indicate fewer errors).
We chose the following 3 D-KEFS tests (Verbal Fluency, Word–Colour Inhibition and the Trail Making Test [TMT]) for use in this study because they are less sensitive to learning and can be used in repeated testing. We chose the Verbal Fluency test to facilitate the comparison of our findings to the literature on noradrenergic-induced deficits because it has face resemblance to word association tests. The Verbal Fluency Test gives participants 60 seconds to name as many words as possible without repeating words more than once. Three-letter fluency conditions are used (F, A and S) and 2 categories are used (animals and boy’s names). The Word–Colour Inhibition Test comprises several parts in which ability to inhibit inappropriate responses while set-shifting is measured. The TMT also assesses participants’ ability to set-shift by drawing trails defined by a series of numbers, the alphabet and switching conditions. For the Verbal Fluency test, the number of words named and errors are recorded; completion times and errors are recorded for the Word–Colour Inhibition test and the TMT.
Following the TSST, participants were once again administered the D-KEFS test, corresponding to samples taken at 10, 20 and 30 minutes after the test.
Diurnal variation of cortisol and α-amylase levels
Salivary cortisol37 and α-amylase38 undergo normal diurnal variation, which is typically dysregulated in conditions such as major depression.39 We asked participants to provide salivary measures at 6 times during the day (upon waking, 30 and 60 min after waking and at 2, 6 and 9 pm) for 2 consecutive days in the middle of their work week as a methodological precaution ensuring the absence of endocrine abnormalities or chronic dysregulation.
Salivary cortisol was analyzed from the saliva samples with a time-resolved fluorescence immunoassay with proven reliability and validity. Salivary α-amylase was quantified by an enzyme kinetic method. Salivettes were analyzed in duplicate in a single assay to avoid confounding related to interassay variability.
Statistical analyses
We performed analyses using the SPSS statistical package version 14 (SPSS Inc.). We used χ2 tests and t tests for sample characterization. We excluded 2 control participants from the CPT analyses who did not understand the CPT instructions (hit proportion < 5%).
Salivary measures
Participants with more than 3 missing values after salivette quantification or 2 or more missing salivettes from the postanticipation period were excluded from TTST salivary analyses. This resulted in the exclusion of 4 relatives and 6 controls from both the salivary α-amylase and cortisol analyses.
We used similar exclusion criteria for the characterization of diurnal variation. In this case, however, participants provided salivettes for 2 days; therefore, the exclusion criteria were applied to individual days of observation. Specifically, a day was excluded if 2 or more salivettes were missing after salivette quantification. This resulted in the exclusion of 15 days of observation (41 d included in the analyses) and the full exclusion of 4 relatives and 3 controls from diurnal cortisol analyses. We excluded 16 days of observation (40 d included in the analyses) and 3 relatives and 3 controls (full exclusion) from the diurnal α-amylase analyses.
We did not exclude any participants from the analyses of neuropsychologic performance regardless of their inclusion in the diurnal or TSST salivary analyses.
The distribution of salivary cortisol and α-amylase levels was verified for skewedness, and we log-transformed the data to approximate a normal distribution.
Diurnal variation and effect of the TSST on salivary α-amylase and cortisol
We used repeated-measures analysis of variance (ANOVA) to analyze the diurnal variation of salivary α-amylase and cortisol. We examined salivary α-amylase and cortisol levels separately. For the analysis of diurnal variation, all 6 salivettes from each day were included as the repeated-measure time. The between-subject effect (group) was included. Final estimates are reported for within-subject time and time × group interaction.
For the analysis of the effect of TSST on salivary amylase and cortisol, all 8 salivettes were included as the repeated-measure stress. The between-subject effect (group) was included. Final estimates are reported for within-subject stress and the stress × group interaction. When we re-evaluated the models to control for history of major depression, the dichotomous variable depression was included as a covariate. Final estimates for within-subject stress, stress × depression and stress × group interactions are reported.
We tested the assumption of sphericity using the Mauchly test of sphericity, and, if the result was significant (p < 0.05), we applied the Huynh–Feldt correction.
Area-under-the-curve analysis
We evaluated the confounding influence of chronic stress (measured by use of the Trier Inventory of Chronic Stress) using the Pearson correlation between measures of stress and area under the curve (AUC) for cortisol and α-amylase. We calculated AUC for the salivettes collected on examination in the laboratory to relate the dynamic variation as a consequence of the TSST. We calculated AUC using well-established trapezoidal methods relative to baseline measures, thus representing the changes from baseline levels (AUCi).40
Effect of the TSST on neuropsychologic performance
We used repeated-measures ANOVA to analyze the effect of the TSST on neuropsychologic performance. Each neuropsychologic test was examined separately. We were primarily interested in errors; we therefore included scaled error scores as the repeated measure. The Verbal Fluency Test, however, does not give standardized scores for errors; therefore, we instead used scaled performance scores as the repeated measure. We included both performance and error indicators in our analyses to account for changes in these related measures in retesting. This approach allowed a within-subject method of controlling for the effect of different completion times between trials on error rates.
We included the before and after TSST error scaled scores (performance for the Verbal Fluency Test) as the repeated measure stress. Group was included as the between-subject effect. Because each neuropsychologic test provides a scaled score for performance and errors, we included a covariate for improvement that corresponded to the difference in performance ([post-TSST minus pre-TSST]/pre-TSST). Final estimates are reported for the within-subject effect stress, stress × improvement and stress × group interaction.
The assumption of sphericity did not apply to these analyses (Mauchly W = 1.00, χ2 = 0.00).
Two neuropsychologic tests used situations of multiple tests conducted on the same instrument. Specifically, the Word–Colour Inhibition Test and the Verbal Fluency Test included 2 components that are of interest for executive function. We applied a Bonferroni correction for multiple testing for these analyses (2p).
When we re-evaluated the models to control for history of major depression, we included the dichotomous variable depression as a covariate, and we re-estimated the model. Final estimates for within-subject stress, stress × improvement, stress × depression and stress × group interactions are reported.
Results
Participants
The mean age of people who had committed suicide was 37.20 (standard deviation [SD] 12.67) years, and they were all white. The relatives’ relationships to the deceased was parent (n = 3), sibling (n = 9) and daughter (n = 2). Because age and sex influence the stress response,41 we strictly matched relatives and controls on these variables (8 women and 6 men in both groups; relatives had a mean age of 44.85 [SD 13.92, range 23–66] years v. healthy controls with a mean age of 44.07 [SD 13.08, range 24–66] years).
Medications taken by the relatives were statins (n = 2), calcium channel blockers (n = 1), biguanides (n = 2), oral contraceptives (n = 2) and esopramazole (n = 1). The medications taken by the controls (n = 14) were levothyroxine sodium (n = 2), β-blocker (n = 1), oral contraceptives (n = 2) and tamsulosin (n = 1).
Evidence suggests that childhood sexual abuse may be related to hyperresponsivity of the physiologic response to psychosocial stress in conditions such as major depression.42 This is particularly relevant in the familial transmission of suicide.43 Although participants were not directly matched on this variable, a comparable number of individuals reported childhood sexual abuse (relatives: n = 3; controls: n = 2; χ12 = 0.55, p = 0.46).
The menstrual cycle has strong influences on salivary cortisol reactivity in response to the TSST.44 The proportion of women who no longer had menstrual cycles was comparable between groups (relatives: n = 4; controls: n = 3; χ2 = 0.25, p = 0.61). Among women with regular menses, a comparable number of days had elapsed since their last menses (relatives: mean 17.00, [SD 9.83] d; controls: mean 17.40 [SD 14.06] d; t7 = 0.04, p = 0.96). Two women in both groups were taking oral contraceptives.
Alcohol and nicotine consumption are also known to affect salivary indicators.18 Participants were reminded not to consume alcohol before coming to the laboratory. Reported habitual consumption did not differ between the groups (relatives: mean 4.10 [SD 5.91] drinks/wk; controls: mean 2.89 [SD 4.06] drinks/wk; t22 = 0.59, p = 0.56). Similarly, tobacco consumption did not differ between groups (relatives: mean 1.50 [SD 3.03] cigarettes/d; controls: mean 2.21 [SD 5.76] cigarettes/d; t26 = 0.41, p = 0.68).
Chronic and perceived stress have been shown to influence the cortisol awakening response.45 We therefore asked participants to report their levels of such stressors using the Trier Inventory for Chronic Stress (TICS).46 No significant group differences were identified on any of the scale’s 10 subscales (p ≥ 0.14, work overload subscale p = 0.097). The correlation between increases in salivary cortisol and salivary α-amylase were examined with respect to total scores on the TICS. When examining salivary cortisol, we did not observe a correlation between cortisol increases in response to the TSST and total TICS score (r = 0.28, p = 0.30). With respect to salivary α-amylase, a trend toward significance was observed when correlating α-amylase increases in response to the TSST and total TICS sore (r = −0.46, p = 0.07).
Psychiatric and baseline neuropsychologic characteristics are presented in Table 1. As expected, relatives were more likely than controls to have had a major depressive episode in their lifetime (odds ratio 17.33, 95% confidence interval [CI] 1.75–171.66, p < 0.01). No other baseline characteristic differentiated suicide relatives and controls.
Baseline psychiatric characteristics and neuropsychologic performance of relatives of people who had committed suicide and healthy controls
Diurnal variation
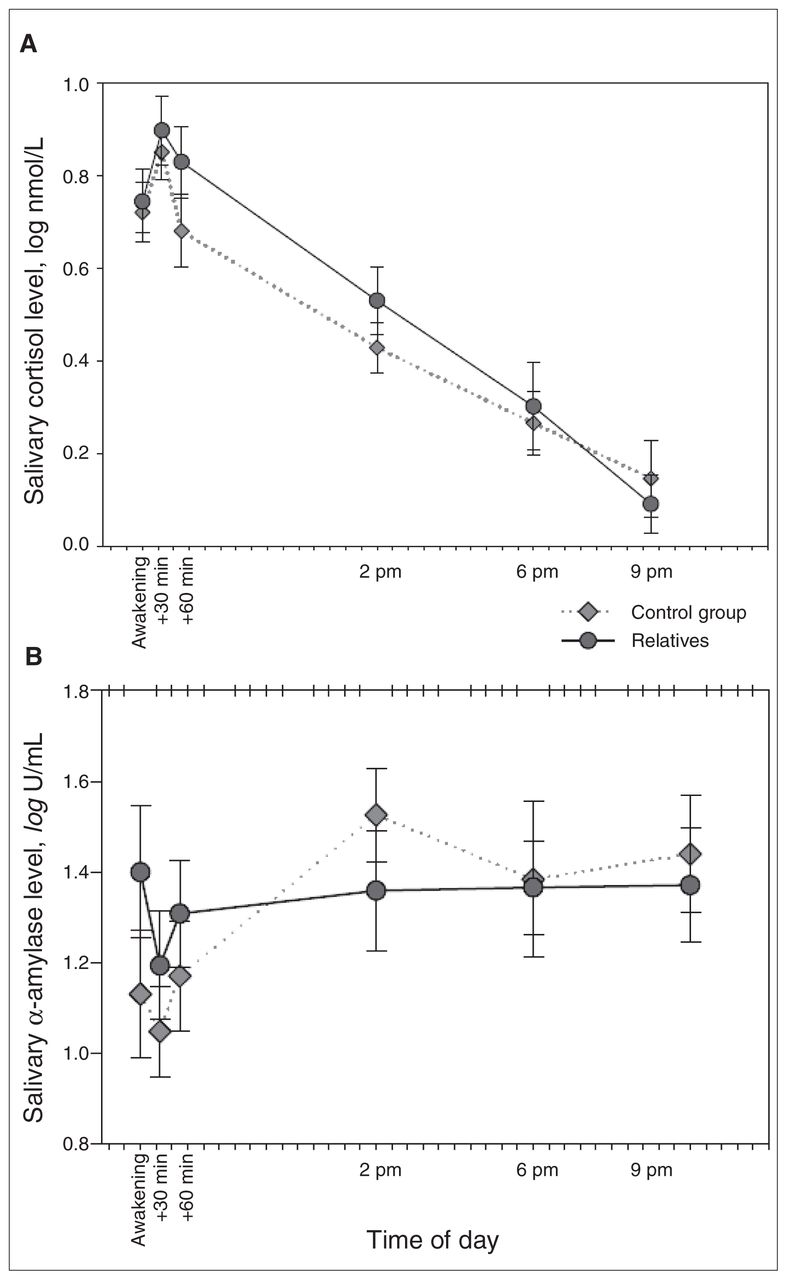
Diurnal variation in cortisol and α-amylase levels are presented in Figure 1.

Daily fluctuations in (A) salivary cortisol and (B) salivary α-amylase. Error bars represent standard errors of the mean.
We first conducted analyses for salivary cortisol diurnal variation using ANOVA. Sphericity was assessed, and it was determined that the data did not meet this assumption (Mauchly W = 0.506, χ142 = 25.28, p = 0.032). Therefore, we applied the Huynh–Feldt correction to the within-subject contrasts. The model revealed a significant effect of time (F4.72,184.42 = 44.09, p < 0.001) but not for the time × group interaction (F4.72,184.42 = 0.59, p = 0.70).
Analyses were then conducted for salivary α-amylase diurnal variation using ANOVA. Sphericity was assessed, and it was determined that the data met this assumption (Mauchly W = 0.636, χ142 = 16.33, p = 0.30). Therefore, we did not apply the Huynh–Feldt correction to the within-subject contrasts. The model revealed a significant effect of time (F5,190 = 4.51, p < 0.001) and a nonsignificant a trend toward a time × group interaction (F5,190 = 1.99, p = 0.08). The relatives had a similar pattern as the controls, but with reduced variability.
Effects of the TSST: salivary analyses
Variation in cortisol and α-amylase in response to the TSST are presented in Figure 2.

Variations in (A) salivary cortisol and (B) salivary α-amylase in response to the Trier Social Stress Test.18 Error bars represent standard errors of the mean.
Analyses were first conducted by use of ANOVA for the effect of the TSST on salivary cortisol. Sphericity was assessed, and it was determined that the data did not meet this assumption (Mauchly W = 0.045, χ272 = 41.64, p = 0.042). Therefore, we applied the Huynh–Feldt correction to the within-subject contrasts. The model revealed a nonsignificant contribution of time (F6.30,100.80 = 1.95, p = 0.08) and a significant effect of the time × group interaction (F6.30,100.80 = 3.07, p = 0.007). More specifically, controls exhibited increases (non–log transformed AUCi, mean 0.77 [SD 7.13] nmol/L), whereas relatives exhibited TSST-induced decreases in cortisol (non–log transformed AUCi, mean −2.46 [SD 5.51] nmol/L) during the post-TSST period.
We then conducted analyses using ANOVA for the effect of the TSST on salivary α-amylase. The data did not meet the assumption of sphericity (Mauchly W = 0.024, χ272 = 50.28, p = 0.005), and we applied the Huynh–Feldt correction to the within-subject contrasts. The model revealed a significant effect of time (F5.84,93.52 = 3.28, p = 0.006) and the time × group interaction (F5.84,93.52 = 2.46, p = 0.031). Controls exhibited elevations in α-amylase in the period following the TSST (non–log transformed AUCi, mean 164.77 [SD 207.52] U/mL), whereas the relatives did not (non–log transformed AUCi, mean 28.42 [SD 170.41] U/mL).
Effects of the TSST: neuropsychology analyses
Statistical models for neuropsychologic performance are presented in Table 2, and significant stress × group interactions are presented in Figure 3.

Effects of the Trier Social Stress Test18 on neuropsychologic performance
The stress × group interaction effect was not significant in the inhibition condition of the Word–Colour Inhibition Test. With respect to the inhibition–switching condition of the Word–Colour Inhibition Test, the model revealed a nonsignificant effect of stress (F1,25 = 0.10, p = 0.92) and significant effects of the stress × improvement interaction (F1,25 = 5.61, p = 0.026) and the stress × group interaction (F1,25 = 5.51, p = 0.027, Bonferroni correction p = 0.054). The relatives did not improve their error rates in the inhibition–switching condition, whereas the controls improved.
With respect to the TMT, the model revealed a nonsignificant effect of stress (F1,25 = 2.97, p = 0.097) and significant interaction effects of stress × improvement (F1,25 = 32.28, p < 0.001) and stress × group (F1,25 = 6.13, p = 0.020). The relatives did not improve their error rates for the switching condition of the TMT.
No significant stress × group effects were observed for the D-KEFS Verbal Fluency test.
Effect of major depression
Owing to expected differences in the prevalence of lifetime major depression between relatives and controls, we performed additional analyses to determine the influence of a history of major depression in these currently euthymic participants. We re-evaluated the ANOVAs to include history of major depression as a dichotomous covariate.
For the effect of the TSST on salivary cortisol, sphericity was again assessed, and the data were not found to meet this assumption (Mauchly W = 0.026, χ272 = 45.58, p = 0.018). We applied the Huynh–Feldt correction to the within-subject contrasts. The model revealed a significant effect of stress (F6.63,99.51 = 2.63, p = 0.017) and the stress × group interaction (F6.63,99.51 = 3.50, p = 0.003) but not the stress × depression interaction (F6.63,99.51 = 1.75, p = 0.11).
For the effect of the TSST on salivary α-amylase, the data were not found to meet the assumption of sphericity (Mauchly W = 0.020, χ272 = 48.46, p = 0.009). Therefore, we applied the Huynh–Feldt correction to the within-subject contrasts. The model revealed a significant effect of stress (F6.35,95.24 = 2.28, p = 0.039) and the stress × group interaction (F6.35,95.24 = 2.31, p = 0.037) but not for the stress × depression interaction (F6.35,95.24 = 1.46, p = 0.20).
Similarly, when the analyses were repeated for the switching condition of the Word–Colour Inhibition test to include history of depression as a dichotomous covariate, the model revealed a nonsignificant effect of stress (F1,24 = 0.02, p = 0.87) and the stress × depression interaction (F1,24 = 0.06, p = 0.80). The model revealed significant effects of the stress × improvement interaction (F1,24 = 5.51, p = 0.032) and the stress × group interaction (F1,24 = 4.38, p = 0.047).
We repeated the analyses for the switching condition of the TMT and included history of depression as a dichotomous covariate. The model revealed nonsignificant effects of stress (F1,24 = 0.26, p = 0.62) and the stress × depression interaction (F1,24 = 0.16, p = 0.69). There was a significant effect for the stress × improvement interaction (F1,24 = 31.16, p < 0.001) and a trend for the stress × group interaction (F1,24 = 3.36, p = 0.08).
Discussion
There is consistent evidence suggesting that suicidal behaviour aggregates in families. Using a high-risk design and a psychosocial-stress paradigm, we investigated the biological and neuropsychologic correlates of the stress response as putative familial traits of suicide risk. Our results indicate that the relatives of people who had committed suicide had blunted stress reactivity for both salivary cortisol and α-amylase in response to the TSST, as well as differences in performance on executive function tests after stress. The effects of suicide vulnerability on the TSST were independent from a previous history of major depression in our participants, all of whom were currently euthymic.
We believe that this study offers a promising direction for suicide research. Despite the small sample size, the robustness of our results and our methodologic stringency suggest that our findings are worth replicating and extending using similar approaches. By choosing heritable indicators and including first-degree relatives, we were able to study the dynamics of a putative component of the etiological chain that may result in suicide when vulnerable individuals are confronted with negative life events.
Salivary measures
Psychologic stress is one of the best-known triggers of the HPA axis. The TSST, one of the best-known stressors to reliably elicit the autonomic nervous system and HPA axis, places the participant in a contextually threatening social–evaluative situation with the perceived possibility of rejection.
Stimulation of the HPA axis via the TSST is associated with a cascade of hormone release. Initially, corticotrophin-releasing hormone is secreted from the hypothalamus, adrenocorticotropin is secreted from the anterior pituitary and, finally, cortisol is secreted from the adrenal cortex.47 α-Amylase has garnered increasing attention as an indicator of physiologic changes following stress. Its secretion is regulated by the sympathetic nervous system48 and is considered a viable indicator of stress response. Much like cortisol, α-amylase exhibits diurnal variation and is stimulated by the TSST.
Contrary to our hypotheses, the differences observed between groups in stress reactivity were related to the relatives’ inability to mount an appropriate stress response. Our hypotheses were guided by postmortem findings suggesting increased adrenal weight49,50 and from dexamethasone suppression test studies suggesting HPA-axis hyperreactivity. One possibility for the difference relates to the presence of active psychopathology, particularly major depression, in both postmortem49,50 and dexamethasone suppression test11–15 studies finding these associations, whereas our relatives were currently euthymic. Major depression is associated with hypercortisolemia and noradrenergic dysfunction, and the long-term negative health effects of metabolic changes accompanying the stress response are consistent with this finding, which may suggest that prolonged and severe environmental insult in the context of major depression may result in suicide. Yet, in our sample, the normal diurnal variation and comparable levels of baseline salivary indicators suggest that relatives are not subject to the long-term metabolic consequences observed in depressed suicide.
The inability to mount a transient stress response, as observed in our euthymic relatives of people who had committed suicide, however, carries different implications than the long-term consequences of stress dysregulation. An appropriate stress response is an adaptive and beneficial physiologic response to stress,47 and with the inability to mount a stress response may come with a range of consequences including executive impairment.
An additional implication of our findings relates to concomitant axis II psychopathology and associated traits. We have recently reported a family study of suicide in which cluster B traits, including levels of impulsive aggression, met full endophenotype criteria for suicide; these traits mediated, in part, the relation between family history of suicide and recurrence among relatives.7 In the context of HPA-axis hypo-reactivity, there have been reports using the dexamethasone suppression test that cluster B personality traits and disorders are related to hypersuppression.51,52 Blunted response to stress and its neurocognitive consequences may represent a co-occurring phenomenon or physiologic consequence of these traits, their familial aggregation and subsequent suicidal behaviour.
Vulnerability to suicide or major depression?
In the current study, the relatives of people who had committed suicide exhibited blunted stress reactivity for both salivary indicators in response to psychosocial stress. At the same time, relatives and controls exhibited comparable diurnal variation. This is important because major depression, posttraumatic stress disorder, chronic fatigue syndrome, fibromyalgia and other conditions characterized by chronic stress and emotional exhaustion42,45,53 are associated with chronic dysregulation of diurnal cortisol variability and blunted stress reactivity. Characterization of diurnal cortisol and α-amylase variation is a strength of the current study, because the normal diurnal variation in these indicators offers a physiologic validation of our eligibility criteria. Instead, this suggests an inability to mount a cortisol response to stress unrelated to active psychopathology among first-degree relatives of people who commit suicide. Moreover, analyses controlling for lifetime history of major depression suggested that the observed differences were related to vulnerability to suicide and not to vulnerability to depression.
Whereas the HPA axis has been previously implicated in depression, this suggests that a family history of suicide is associated with heritable neuroendocrine alterations outside of vulnerability to depression.
Neuropsychology
Executive function tests require the integration of several cognitive functions, which are themselves quite complex. Disruption of any of these functions, including selective attention, abstract thought and cognitive shift, can lead to impaired performance. The executive function tests chosen for the current study are considered heritable,54,55 and therefore appropriate in the context of a family study of suicide.
Despite comparable abstract reasoning and attention, as well as comparable baseline performance on all 3 D-KEFS tests, relatives’ and controls’ performance on the Word–Colour Inhibition Test and Trail Making Test were differentially affected by a controlled laboratory psychosocial stressor. This is the first study, to our knowledge, to examine the effects of a controlled acute psychosocial stressor on executive function performance in this population. More specifically, despite similar baseline neuropsychologic performance, the relatives of people who had committed suicide failed to improve their on the Word–Colour switching/inhibition and TMT switching conditions, an effect independent of previous depression.
Executive function provides neuropsychologic evidence of cognitive flexibility. The TSST induction of changes in performance on executive function tasks improves our understanding of cognitive changes that take place following negative life events that may act as risk factors for suicidal behaviour. Pursuing behaviours resulting in stress generation or mal-adaptive coping strategies consequent to life events may play an important role in the etiology of suicide. Our study suggests that acute psychosocial stress-induced cognitive inflexibility and decreased ability to inhibit inappropriate actions is a potential factor that increases risk for suicidal behaviour.
Limitations
We used single indicators of both the sympathetic nervous system and HPA-axis activation. Future studies should include additional measures, such as heart rate and blood pressure. This study had a small sample size, and therefore larger samples should be investigated and additional neuropsychologic changes characterized. In addition, a larger sample size would allow an investigation of the role of childhood sexual abuse on neuroendocrine reactivity in this population.
Whereas we statistically controlled for the effect of a history of major depression, this is an important confounding variable that should be addressed in future studies of psychosocial stress in this population. At the same time, we recruited relatives without personal histories of suicidal behaviour to maximize sample homogeneity and avoid the confounding influence of previous suicide attempts. However, one could instead speculate that we recruited a group of relatives who possess additional protective effects. Future studies, therefore, should include suicide attempters with a positive family history.
The influence of the traumatic loss of a close relative to suicide on the characteristics of the stress response cannot be ignored, nor can its role be disentangled using the current design. Additional studies using adoption studies would be well suited to addressing this limitation.
Conclusion
The relatives of people who commit suicide exhibit dysregulated autonomic nervous system and HPA-axis activation as well as changes in executive function in response to psychosocial stress. Secondary analyses indicate that this is not related to predisposition to major depression.
Acknowledgements
This study was supported in part by the Canadian Institute of Health Research (MOP 57924). Dr. Turecki is a Fonds de la recherché en santé chercheur boursier.
Footnotes
Previously published at www.jpn.ca
Competing interests: None declared for Mr. McGirr and Ms. Cabot and Drs. Diaconu, Pruessner, Sablé and Turecki. Dr. Berlim has received a grant from the National Alliance for Research on Schizophrenia and Depression.
Contributors: Drs. Turecki and Sablé and Mr. McGirr were involved in the conception and design of the study. Mr. McGirr, Ms. Cabot, and Dr. Diaconu acquired the data, which was analyzed by Drs. Diaconu, Pruessner and Berlim. Drs. Diaconu and Pruessner and Mr. McGirr wrote the manuscript, which was revised by Drs. Sablé, Turecki and Berlim and Ms. Cabot. All authors approved the final version submitted for publication.
- Received September 17, 2009.
- Revision received January 31, 2010.
- Accepted March 30, 2010.
References
In this issue

{kind=link}
{kind=link}
{kind=link}
Article tools





