





Abstract
It has been suggested that bilateral symmetry may impose a cost for animals relying on camouflage because symmetric color patterns might increase the risk of detection. We tested the effect of symmetry on crypsis, carrying out a predation experiment with great tits (Parus major) and black-and-white–patterned, artificial prey items and background. First, we found that detection time was significantly longer for a highly cryptic, asymmetric pattern based on a random sample of the background than for its symmetric variants. Second, we were able to arrange the elements of a prey pattern in a way that the resulting asymmetric pattern was highly cryptic and, furthermore, its symmetric variant was highly cryptic as well. We conclude that symmetry may impose a substantial cost on cryptic patterns, but this cost varies among patterns. This suggests that for prey, which predators typically view from an angle exposing their symmetry, selection for pattern asymmetry may be less important and selection for decreased detectability cost of symmetry may be more important than previously thought. This may help to understand the existence of so many prey with cryptic, symmetric color patterns.
Cryptic color pattern or camouflage is an adaptation that prey use to decrease their risk of becoming detected by visual predators (Cott, 1940; Edmunds, 1974; Merilaita et al., 1999; Ruxton et al., 2004). To achieve a low risk of detection by means of color pattern, it is important that the color pattern of the prey should not stand out from the visual background. Therefore, visual resemblance in colors and pattern geometry between the prey and its background decreases the risk of detection (Cott, 1940; Endler, 1978; Merilaita and Lind, 2005; Merilaita et al., 1999; Norris and Lowe, 1964). Also, because the body of a prey or some parts of it may have a characteristic shape that helps predators to detect and recognize the prey, it may be important to make the detection of prey shape more difficult. The use of colors and pattern geometry to obscure prey shape and contour is known as disruptive coloration (Cott, 1940; Edmunds, 1974; Thayer, 1909). For example, it has been suggested that marginal pattern elements that touch the outline may be used to break up the continuity of the outline and distractive marks may be used to draw the attention of the observer from the outline and other characteristics that facilitate recognition (Cott, 1940; Cuthill et al., 2005; Merilaita, 1998; Merilaita and Lind, 2005).
The majority of taxa belonging to the Animal Kingdom are bilaterally symmetrical. Consequently, predators may encounter and detect prey, especially those that are dorsoventrally flattened, typically from an angle that reveals their symmetry. Symmetry perception is widespread among animal taxa, and for example, bees have been shown to be able to detect and generalize symmetry as an attribute (e.g., Giurfa et al., 1996, and references therein). It has been suggested that symmetry detection may be useful for predators searching for prey and, consequently, color pattern symmetry may be detrimental or even a major failing for prey relying on camouflage as symmetry may increase the risk of detection (Forsman and Merilaita, 1999, 2003; Kirkpatrick and Rosenthal, 1994; Osorio, 1994). This is in agreement with experiments demonstrating that symmetry can increase the efficacy of antipredator warning color patterns (Forsman and Herrström, 2004; Forsman and Merilaita, 1999).
On the other hand, it is highly exceptional that an animal with a symmetric body has a highly asymmetric color pattern. Even prey with cryptic coloration do as a rule have symmetric color patterns. However, this may be due to developmental or genetic coupling between color pattern and other traits (Forsman and Merilaita, 2003). Thus, the rarity of cryptic prey with asymmetric color pattern does not necessarily mean that symmetry would be unimportant for natural selection on cryptic color patterns. However, so far, the effect of color pattern symmetry on prey detection has not been empirically tested.
It is important to investigate the costs associated to antipredator coloration because of the significance of such costs both for the choice of defense strategy (e.g., between camouflage and warning coloration) and for the selection on the appearance of prey coloration (Merilaita and Tullberg, 2005; Ruxton et al., 2004; Speed and Ruxton, 2005). Therefore, to better understand antipredation coloration, it should be of interest, first, to experimentally study whether symmetry does indeed incur a cost for crypsis and, second, to investigate the effects of symmetry on selection on cryptic color patterns. In this paper, we have studied these questions, focusing on two main objectives. First, by comparing the detection times of an asymmetric pattern and its two bilaterally symmetric variants, we experimentally show that symmetry does in fact incur a potential detectability cost for cryptic color patterns. Second, we wanted to find out whether the cost of symmetry is invariable or if it is possible to decrease this cost. Therefore, our aim was to produce another cryptic and asymmetric pattern, but now, in comparison to the first pattern, with a low detectability cost when transformed to a bilaterally symmetric pattern. In addition, we will critically scrutinize the idea that cryptic coloration is maximized by a coloration that represents a random sample of the background (Endler, 1978, 1984, 1991; but see Merilaita and Lind, 2005).
METHODS
Predators, backgrounds, and prey items
To conduct this study, we created artificial prey items and backgrounds and trained great tits (Parus major) that we used as predators to search for the prey items. Methods of this study are very similar to the methods of Merilaita and Lind (2005). We trapped birds using mist nets and performed the experiment at Tovetorp Zoological Research Station in southeast Sweden. The study was performed with permission from the Swedish Ethical Board for conducting behavioral research. All birds were housed indoors separately in holding cages (80 × 60 × 40 cm) where they were provided with water, suet, and sunflower seeds ad libitum. Prior to this experiment, all birds were used in another experiment (Merilaita and Lind 2005). However, because the prey and background patterns were completely different in the two experiments and the birds were retrained before the second experiment, we have no reason to expect that the first experiment would have interfered with the results of the second experiment reported here. Further, all birds were subjected to the same treatment in the previous experiment.
We chose the prey and background patterns for the present experiment so that the detection task would be difficult enough to introduce variation in detection time between prey with different patterns, but not too difficult, so that the great tits would not lose their motivation to search for the prey. We used artificial, black-and-white–patterned prey and background. To avoid variation among background sheets and prey items, we used the same copying machine with the same toner and paper throughout the experiment when reproducing backgrounds and prey from the printed originals.
We created an A4-sized background (20 × 29 cm), using Corel Draw 10 (Corel Corp., Ontario, Canada). The pattern is shown in Figure 1a. To make background boards for the experiment, we glued the printed backgrounds to equally sized pieces of corrugated cardboard, using solvent-free glue stick (Scotch, 3M, Cergy, France). On each board, we made one hole in a random location, put a peanut chip in the hole, and covered the hole by lightly gluing a prey item on it. Thus, the predators had to search for the prey items on the backgrounds to find the peanut chips.
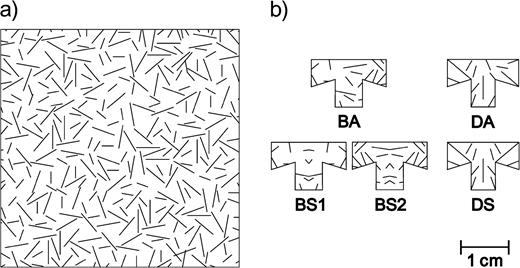
A sample of the background (a) and the five prey color patterns (b) used in the experiment. There were two asymmetric patterns, a background-matching pattern, BA, and an arbitrarily made disruptive pattern, DA. Three symmetric patterns were created from the asymmetric ones, BS1 and BS2 from BA and DS from DA.
All prey items had an identical shape (Figure 1b). We used Corel Draw 10 (Corel Corp., Ontario, Canada) to create the prey patterns. To achieve the main objectives of this study, that is, to demonstrate (1) that there is potentially a substantial cost associated with color pattern symmetry in cryptic prey but (2) that not all cryptic patterns must suffer equally much from such cost, we created two prey sets. The sets consisted of an asymmetric pattern and one or two bilaterally symmetric transformations of the asymmetric pattern. In the first prey set, crypsis was achieved through background matching (Endler, 1978; Merilaita and Lind, 2005; Merilaita et al., 1999; Norris and Lowe, 1964). Therefore, the asymmetric pattern represented a sample of the background. To ensure that the sample would provide a highly cryptic pattern (cf. Merilaita and Lind, 2005), we first took 20 samples of the background by randomly choosing the location and the orientation for the prey outline on the background with respect to the x- (0–21 cm) and y-coordinates (0–29 cm) and the rotation (1°–360°). Then, based on preliminary testing using our own judgment (i.e., testing how difficult the prey patterns were for us to detect on the background) and trials similar to the experiment, using three birds that were excluded from the experiment, we chose the pattern that appeared most cryptic. We made two bilaterally symmetric patterns (BS1 and BS2), based on the lateral halves of the asymmetric pattern (BA). These patterns are shown in Figure 1b. With this first set of patterns, we wanted to demonstrate the potential cost of symmetry on a cryptic pattern.
With the second prey set, we wanted to find out if it is possible to decrease the cost of symmetry. We created asymmetric patterns arbitrarily with the aim that they would not only resemble the background but also disrupt the prey shape (Cott, 1940; Cuthill et al., 2005; Merilaita, 1998). To achieve the disruptive effect, we used the lines as false outlines to break down the prey shape as well as to change (divide) some angles. We created 10 different patterns and, again, chose one highly cryptic pattern with the help of our own judgment and trials with the three birds. However, because this time we wanted the bilaterally symmetric transformation of the chosen pattern as well to be highly cryptic, we included the symmetric transformations in the preliminary testing. The second set consisted of an asymmetric pattern (DA) and its bilaterally symmetric transformation (DS), shown in Figure 1b.
Training procedure
As mentioned above, all birds took part in another experiment before they were used in the present study. For the previous experiment, we trained all birds to associate the artificial prey items with food. This is described in detail in Merilaita and Lind (2005). In the previous experiment, the birds searched for artificial prey with the same shape but with different patterns and on a different background compared to the present experiment. Therefore, it was enough for this experiment to train the birds to recognize the prey patterns. For this purpose, all birds went through one step of training. We conducted the training in the experimental cages. As food we used peanut chips (approximately 2 × 2 × 2 mm). The background was first glued to a piece of cardboard (15 × 21 cm), and then the five prey types were glued lightly (from three points) to cover small holes in the background board, each containing a peanut chip. We folded one “wing” of the “T” of one randomly chosen prey item to make it easier to detect and hence stimulate the birds to search for more prey on the background.
Experimental procedure
The experiment was carried out in plywood cages (55 × 90 × 70 cm). Observations were made through an opening (10 × 12 cm), covered with a mesh net, at the top of the front door. The birds had always free access to fresh water. At the bottom of the right side of the cage, we made an opening (2 × 32 cm) where we could insert training boards and experimental background boards. When not used, this opening was covered by a lid. A perch was located underneath the ceiling opposite this opening, and cages were lit by two lamps (Philips, Eindhoven, the Netherlands; fluorescent, 9 W). We kept the experimental room dark so as not to disturb the birds while observing them. Altogether, we used 19 trained birds in this experiment.
The birds were acclimatized to the experimental cage for 45–60 min before the start of the experiment. During the acclimatization, no food was available so that they would be motivated to search for food at the start of the experiment. During the experiment, all the five different prey types were presented three times to each bird in sequence. Hence, each bird searched for 15 prey items in total. We randomized the order of presentation of the five different prey types but balanced the design so that all prey types would be presented equally many times as the first prey. When the experiment was completed, the bird was released at the trapping site.
We used effective search time, measured by a stopwatch, as the measure of the probability of detection. We defined effective search time as the time a bird spent on the background board searching for prey. If a bird did not find a prey on a background board within 600 s, either because the prey was difficult to find or because of low motivation, the board was taken out of the cage and presented again after the remaining boards had been presented. To avoid confusing the lack of motivation with low probability of detection, we used the effective search time measured in the latter presentation (when the prey was found within 600 s) in the analysis. Five birds did not find one prey within the first 600 s, and five birds did not find two prey during the first 600 s.
Statistical analysis
For each bird and prey type, we calculated the mean effective search time. These values were log transformed to achieve normally distributed data (Kolmogorov-Smirnov test for normality: p > .66 for all variables after the transformation). Because each bird looked for all the five prey types, we used paired t tests in the comparisons. To avoid committing type-I errors due to multiple comparisons, we used the Dunn-Šidák method to adjust α-levels (Sokal and Rohlf, 1995). Only the corrected p values are presented. Statistical tests were performed using SPSS 11 (SPSS Inc., Chicago, IL).
RESULTS
We found that for the first prey set, in which the pattern was based on a random sample of the background, the symmetry transformation markedly affected the probability of detection. Thus, compared to the asymmetric pattern, BA, the effective search times of the two bilaterally symmetric variants were significantly shorter (BS1: t18 = 3.35, p = .04; BS2: t18 = 3.77, p = .001; Figure 2). The effective search times of the two symmetric patterns, BS1 and BS2, did not differ significantly (t18 = 0.95, p = .36).
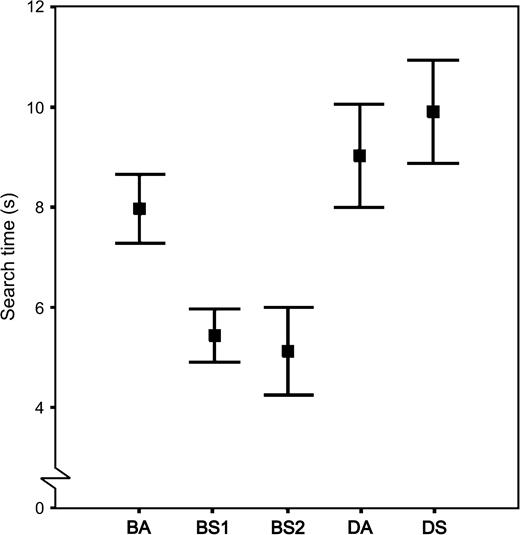
Mean search times (±SE) in seconds for the five different prey color patterns.
On the other hand, the effective search times did not differ significantly between the asymmetric and the symmetric patterns of the second prey set, DA and DS (t18 = 0.99, p = .34; Figure 2). Yet, the effective search times of both the patterns in the second set were relatively long. This was shown because they did not significantly differ from the search time of background-matching, asymmetric pattern of the first set (DA: t18 = 0.89, p = .39; DS: t18 = 2.06, p = .054; Figure 2). Also, the effective search times were significantly longer for both the disruptive patterns than for the left side–based symmetric, background-matching pattern, BS1 (DS: t18 = 4.70, p < .001; DA: t18 = 5.44, p < .001; Figure 2), and the right side–based symmetric, background-matching pattern, BS2 (DS: t18 = 7.50, p < .001; DA: t18 = 4.24, p < .001; Figure 2). In other words, we could not detect a cost associated with the symmetry transformation for the second prey set, indicating that we had been successful in our attempt to decrease the symmetry cost in a relatively highly cryptic pattern.
DISCUSSION
There are two main findings in this study. First, for the first prey set, the transformation of the asymmetric prey pattern to bilaterally symmetric patterns decreased prey crypsis. In this set the detection time was shorter for the two symmetric patterns, BS1 and BS2, than for the prey with the asymmetric pattern, BA. This confirms the suggestion that there is a potential cost to cryptic prey for bilateral symmetry (cf. Forsman and Merilaita, 1999; Osorio, 1994). The second main result is that, in contrast to the first prey set, in the case of the second prey set we did not find a difference in the detection time between the asymmetric prey, DA, and its symmetric transformation, DS. This indicates that the cost in terms of increased detectability due to pattern symmetry may vary markedly among different symmetric patterns.
Our results have several implications for the evolution of cryptic coloration, specifically in prey that are typically seen by predators from an angle that reveals their symmetry. First, the results suggest that although color pattern symmetry may impose a cost in terms of decreased crypsis, not all bilaterally symmetric, cryptic patterns carry an equal cost for symmetry. It is even possible that for some symmetric patterns there may be no cost whatsoever. Such variation in the cost due to pattern symmetry suggests that selection against pattern symmetry may not be as important as previously expected (cf. Forsman and Merilaita, 1999, 2003; Kirkpatrick and Rosenthal, 1994; Osorio, 1994). Instead of selection for pattern asymmetry, this emphasizes the importance of selection for a decreased detectability cost due to symmetry, when considering the appearance of cryptic patterns. Further, such possibility to decrease the detectability cost due to pattern symmetry may also help us to understand the existence of so many prey with cryptic, symmetric color patterns.
Another implication from our results deals with background matching. It has been suggested that coloration that represents a random sample of the background maximizes crypsis (Endler, 1978, 1984, 1991). In a recent study we showed that the generality of this idea is limited because such random sampling is neither sufficient nor necessary for the minimization of the probability of detection (Merilaita and Lind, 2005). The results of the present study suggest another argument against that idea, when it comes to bilaterally symmetric patterns. Our study shows the importance of how pattern elements are distributed on a prey color pattern in relation to the symmetry axis (see subsequently). Because of the variation among color patterns in the cost of symmetry, it is unlikely that color pattern based on any random sample of a background would maximize crypsis. Instead, we should expect that only a subsample of all random samples of a background would provide a decreased cost of symmetry. Further, the color pattern with minimal cost of symmetry need not necessarily be a pattern that matches a sample of the background.
Then, why does symmetry influence the probability of detection? A possible explanation is as follows. Perceptual organization that results in decision of which regions in the visual field correspond to objects and which correspond to background is called figure-ground segmentation. One factor that the visual system uses in figure-ground segmentation is symmetry: all else being equal, symmetrical regions tend to be perceived as figure (e.g., Driver and Baylis, 1996; Palmer, 1999). The edge between a figure and ground is assigned as the one-sided contour of the figure, and attention tends to be drawn to figure (Baylis and Driver, 1995; Driver and Baylis, 1996; Palmer, 1999). Consequently, symmetry can enhance detection of cryptic prey because of these perceptional mechanisms.
Our study suggests something about why the symmetric patterns based on random samples of the background, BS1 and BS2, were easier to detect than the symmetric, disruptive pattern, DS, although it was not designed specifically to answer that question. One difference between these two prey categories was that for the randomly sampled patterns the symmetry transformation created by the axis of symmetry merged pattern elements deviating from the pattern elements on the background (i.e., some straight lines were turned into chevrons). In the DS pattern such deviating elements were absent. It is possible that these deviating elements increased the risk of detection due to decreased match between the prey and the background. Note, however, that not only the BS prey but probably all the prey types suffered to some extent from deviating elements partly because the prey partially covered some elements on the background and also partly because merging of pattern elements of the prey and the background may have produced deviating elements. Another possible explanation, not necessarily mutually exclusive with the first one, is also related to the elements by the symmetry axis, namely, symmetries near the symmetry axis may be especially useful in symmetry detection. This is suggested by studies on humans showing that symmetry detection employs a limited spatial region over which subjects integrate information (e.g., Dakin and Herbert, 1998). Accordingly, symmetric or symmetrically located elements at the symmetry axis of a prey (such as the chevrons) may be especially revealing, facilitating prey detection.
In sum, our study shows that there is a potential cost due to symmetry for cryptic prey in terms of increased risk of detection. However, the study also shows that this cost may significantly vary among different color patterns. Therefore, we expect there to be natural selection for cryptic patterns that minimize the cost due to symmetry. Our study suggests some predictions that could be tested by analyzing color patterns of real prey relying on camouflage. One of the predictions is that, in contrast to the idea of crypsis being maximized by a random visual sample of the background, we would expect selection for background-matching select against distinct pattern elements at the symmetry axis that would probably be created by a simple symmetry transformation of a sample of a background. Another prediction is that selection for disruptive coloration may result in cryptic, symmetric patterns with less symmetric arrangements of pattern elements near the symmetry axis than further away from it.
We would like to thank Anders Forsman and Will Cresswell for valuable comments. This study was supported by the Swedish Research Council (S.M.).
References
Baylis GC, Driver J,
Cuthill IC, Stevens M, Sheppard J, Maddocks T, Párraga CA, Troscianko TS,
Dakin SC, Herbert AM,
Driver J, Baylis GC,
Endler JA,
Endler JA,
Forsman A, Herrström J,
Forsman A, Merilaita S,
Forsman A, Merilaita S,
Merilaita S,
Merilaita S, Lind J,
Merilaita S, Tullberg BS,
Merilaita S, Tuomi J, Jormalainen V,
Norris KS, Lowe CH,
Ruxton GD, Sherratt TM, Speed MP,
Speed MP, Ruxton GD,

{kind=link}
{kind=link}