
Abstract
Memory for contextual information and target-context integration are crucial for successful episodic memory formation and are impaired in patients with Korsakoff’s syndrome. In this paper we review the evidence for the notion that a context memory deficit makes an important contribution to the amnesia in these patients. First, we focus on anterograde memory for contextual (spatial and temporal) information. Next, the use of contextual cues in memory retrieval is examined and their role in retrograde amnesia and confabulation. Evidence on the role of contextual cues and associations in working memory is discussed in relation to the underlying neurocognitive mechanisms and their dissociation from long-term encoding. Finally, we focus on implicit learning of contextual information in Korsakoff patients. It can be concluded that Korsakoff patients are impaired in the explicit processing of contextual information and in target-context binding, both in long-term (retrograde and anterograde) memory and in working memory. These results extend the context memory deficit hypothesis. In contrast, implicit contextual learning is relatively preserved in these patients. These findings are discussed in relation to evidence of dysfunction of the extended diencephalic-hippocampal memory circuit in Korsakoff’s syndrome.
Similar content being viewed by others


Introduction
When remembering events from our past or storing new information (often referred to as target information), contextual processing is a crucial aspect. That is, everyday, episodic memory becomes meaningless without a context: the “where and when” of our past, present and future experiences. While there is agreement that target memory (the to-be-remembered information itself) and context memory can be distinguished, there is less consensus about the definition of the concept of “context” within the memory literature. Rudy’s (2009) defines context as “a set of concurrent independent component features that potentially can be sampled by an individual” (Rudy 2009, p. 573). Furthermore, he distinguishes two defining properties of features that make up a context. First, these features and their interrelationship must be stable (i.e., independent from the observer). Second, while stable, the constellation is not fixed, as it can be rearranged or manipulated (changing the context). In this view, context is regarded as “background information” that can be an actual environment (for instance, the room in which an experiment takes place), the colour or font of words that are presented on a computer screen, the location of objects on a monitor or the order in which events take place. This spatial or temporal-order background information in itself is meaningless, unless a link or association can be established with the target information, such as the words, objects or experimental setting. These target-context associations form the basis of episodic memory. In addition, contextual information may serve as a retrieval cue. That is, during recall, contextual information may facilitate the memory retrieval through its association or interaction with the target information during the learning phase (see, e.g., Tulving 1983).
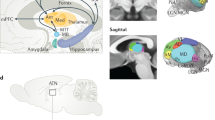
Within the cognitive system, domain-specific processing takes place in specialised modules that often have a distinct neural substrate. For example, the fusiform face area subserves the perception of faces, the semantic network processes verbal information, and colours and motion are processed within V4 and V5 respectively. In order to form a coherent episode, information coming from these different neurocognitive modules has to be integrated or bound. A neuroanatomical account of long-term episodic memory formation, suggests that to-be-remembered information from the cortical association areas is processed via the perirhinal cortex (object information), and the parahippocampus (spatial information) and integrated or bound within the entorhinal cortex (Squire et al. 2004). Subsequently, the actual storage may take place in the hippocampus (Squire et al. 2004), or it may act as index or pointer to where those memories are stored (Nadel and Moscovitch 1997). The hippocampi are connected with the diencephalon via the fornices that end in the mammillary bodies, which in turn project to the thalami (see Fig. 1 for a schematic overview of this functional model). This hippocampal-diencephalic circuitry is often regarded as the “extended hippocampal system” (Aggleton and Brown 1999) that is crucial for the formation of new memories. Lesions in this extended memory system do indeed produce memory deficits.

A functional overview of the extended hippocampal-diencephalic memory system. Nonintegrated input from the association areas (such as spatial or object information) is processed in the parahippocampal and perirhinal cortices and then integrated or bound as an “episode” in the entorhinal cortices. Storage then takes place through the hippocampi that are connected via the fornices to the mammillary bodies. Subsequently the (anterior) thalami project to the neocortex where the episodic information is permanently represented
In Korsakoff’s syndrome, the profound amnesia is directly related to bilateral lesions in the diencephalon (notably the thalamus and mammillary bodies), but perhaps also to the hippocampus proper (Visser et al. 1999; Sullivan and Marsh 2003; Sullivan and Pfefferbaum 2009, but see Colchester et al. 2001). It has been argued that the amnesia in Korsakoff patients is a direct result of the inability to remember contextual information (Mayes et al. 1985). Others have suggested that a ‘binding deficit’ (affecting contextual memories) may underlie the profound amnesia in Korsakoff patients (see for a discussion e.g., Chalfonte et al. 1996). Moreover, in forms of memory that are not part of the episodic memory system, contextual processing may also be important: for example during working memory tasks in which associations have to be held active for a brief period of time (e.g., Piekema et al. 2010) or in implicit learning of visuospatial patterns that do not require conscious recall (see, for instance, Chun and Jiang 2003) or complex visuospatial motor sequences (e.g., Witt and Willingham 2006). However, the role of dysfunction of the diencephalic-hippocampal memory system in these nonepisodic forms of memory and learning is less clear. In this paper, studies of context memory in Korsakoff patients will be reviewed, focusing on the role of context in anterograde amnesia, contextual cues in retrograde (autobiographical) amnesia, contextual working memory and contextual cues in implicit learning, as well as the implicit acquisition of contextual information.
Anterograde Amnesia in Korsakoff’s Syndrome
The Origins of the Context Memory Hypothesis
Korsakoff (1889) himself emphasised the difficulty of his patients in aspects of context memory, particularly temporal sequence. However, it was Huppert and Piercy (1976, 1978) who first proposed the context memory deficit hypothesis in the modern, empirical literature. Huppert and Piercy (1976) showed pictures of scenes to eight Korsakoff patients and eight alcoholic controls 24 h before carrying out a recognition memory test (i.e., presenting the previously shown pictures among novel distractor items). On the second day, the participants were shown further pictures of scenes, some of which had been seen the day before, and some of which had not; a Yes-No recognition memory test asked participants to identify as familiar only those pictures seen on the same day. What was found was a highly significant interaction effect between groups and the familiarity of the material, indicating a much stronger tendency among the Korsakoff patients than the controls to say “Yes” to familiar as opposed to unfamiliar pictures, i.e. which day they had been seen on, giving a high rate of false positives. It appeared that the Korsakoff patients had severe difficulty in discriminating between items seen 10 min previously and items seen 24 h previously, but healthy controls did not have difficulty in making this discrimination. The Korsakoff patients performed well in making judgements of familiarity per se, suggesting that the Korsakoff patients remembered the attributes of an item far more efficiently than information concerning the context in which the item was presented. The authors postulated a selective deficit in context memory, such that context memory is much more vulnerable than memory for item-familiarity in Korsakoff patients.
Huppert and Piercy (1978) tested this hypothesis more rigorously by carrying out an experiment across 2 days, in which they varied not only the occasion of presentation of the stimuli (pictures of scenes), but the frequency with which each stimulus was presented (once or 3 times on each of Day 1 and Day 2). Retention testing began 10 min after presentation on Day 2. Participants had to say whether each picture had been seen “today” or “yesterday”, and whether they had seen the picture “once” or “3 times”. In both controls and eight Korsakoff patients, recency judgements were influenced by the frequency of presentation, and frequency judgements by the recency of presentation. However, healthy controls were significantly better able to discriminate between recency and frequency than were Korsakoff patients, indicating that the controls’ decisions were influenced jointly by trace strength and specific information about the time and frequency of presentation. By contrast, the Korsakoff patients were unable to make recency/frequency discriminations, suggesting that their judgements were determined solely by trace strength, and not by specific contextual information. As an earlier experiment (Huppert and Piercy 1977) had shown that patients’ recognition memory reflected the total presentation time during which they were exposed to the stimuli (Korsakoff patients performed as well as controls on a recognition memory test, provided that the stimuli was shown for approximately 4 times as long), the authors concluded that the contextual memory deficit operates during the initial acquisition of information.
At much the same time, Winocur and Kinsbourne (1978) carried out an experiment in which Korsakoff patients performed paired-associate learning tasks in experiments in which the context at learning and retrieval was varied. The authors found that strong environmental cues present at both learning and retrieval benefitted the Korsakoff patients’ performance. They argued that the Korsakoff patients have a defect in (spontaneous) context retrieval, which was made good where there were strong environmental cues. In other words, they were also identifying a deficit in an aspect in contextual memory, but placing the deficit at retrieval, rather than at encoding.
The Mayes and Meudell Context Memory Deficit Hypothesis
Meudell and Mayes followed up these investigations. These authors had previously carried out a series of studies showing that Korsakoff’s patients amnesic deficit could not be attributed to either a specific failure in semantic encoding, as advocated by Butters and Cermak (1980), or to a specific retrieval deficit and vulnerability to interference effects, as advocated by Warrington and Weiskrantz (1970). In most of these experiments, Meudell and Mayes (1982) employed ‘matching procedures’ (by varying the exposure times to which participants were shown the stimuli, or the delays until testing, such that controls had prolonged delays) so that performance levels were equated in the two groups (amnesic, control) of participants. The purpose of this was to check that differences between the groups across orienting (encoding) tasks or retrieval tasks was not an artefact of differences in memory strength.
In their investigations of context memory, Meudell and Mayes adopted a similar approach. Meudell et al. (1985) examined recency and frequency judgements in Korsakoff patients and in healthy controls in a similar manner to that employed by Huppert and Piercy (1978). However, in their study, the healthy controls were tested at prolonged delays, such that their overall level of recognition memory performance was matched as possible to that of the Korsakoff patients. Meudell et al. (1985) found that the amnesic patients did indeed confuse recency and frequency judgements as Huppert and Piercy (1978) had found. By contrast, the healthy participants could still make memory judgements about temporal sequence, independently of the frequency of exposure to the stimuli. This allowed Meudell et al. (1985) to identify a deficit in temporal context memory in their Korsakoff group. Somewhat similarly, Shoqeirat and Mayes (1991) showed that, on a task originally developed by Smith and Milner (1981), amnesic patients were disproportionately impaired in making spatial location judgements, compared with healthy controls, even though the two groups had been matched in terms of the ‘target’ recognition memory by testing the healthy participants at prolonged delays.
As a result of these and other investigations, Mayes and colleagues (Mayes et al. 1985; Mayes 1988) postulated that a context memory impairment might be the fundamental deficit in amnesia. They argued that Korsakoff patients suffer from a selective inability to remember background context, i.e. spatio-temporal or intrinsic context. However, they emphasised the need to control for memory strength in examining for specific contextual memory deficits, and also to control for what might be incidental consequences of frontal cortical damage, rather than essential aspects of the core memory deficit. Subsequent investigations by Mayes and his group conceptualised the context memory deficit as a component of a more generalised impairment in binding complex associations (Mayes and Downes 1997), and later as an important aspect in a failure of recollective memory, attributable to pathology within the extended hippocampal circuitry (Montaldi and Mayes 2010).
Other Studies on Spatial and Temporal Context Memory
Various researchers have investigated this topic further, generally finding specific deficits in context memory. Kopelman (1989) examined (i) a retrospective dating of famous news events, (ii) an anterograde temporal context memory task, in which subjects had to decide which of two series pictures had appeared in, separated by approximately 45 min, and (iii) an orientation questionnaire. Korsakoff patients showed a U-shaped curve (i.e. both a primacy and recency effect) in dating famous news events. They were significantly impaired, compared with healthy controls, but the degree of their impairment was unrelated to whether or not they showed superimposed frontal cortical atrophy on CT brain scans. On the anterograde temporal context test, recognition memory was matched to that of healthy controls, and the Korsakoff patients showed disproportionate and statistically significant impairment for temporal context, which was unrelated to the presence or absence of frontal atrophy. Likewise, on the orientation questionnaire, Korsakoff patients were again significantly impaired, unrelated to the degree of frontal atrophy. These three temporal context tasks correlated significantly together, and also with the severity of the patients’ memory overall impairment (as indicated on the Wechsler scales), but only weakly with performance on a series of measures of executive function.
Parkin et al. (1990a) compared 10 diencephalic amnesic patients, 15 patients with temporal lobe pathology, and healthy controls on a task where previously viewed stimuli subsequently became ‘distractors’ and distractors became targets. On this task, Korsakoff patients had much more difficulty in making temporal context/recency judgments than did either the healthy volunteers or patients with temporal lobe pathology (see also Hunkin et al. 1994). In contrast, these authors argued that medial temporal pathology was more damaging to spatial context memory.
Similarly, Chalfonte et al. (1996) found spatial context impairment in a group of mixed-aetiology amnesic patients, which was attributed to medial temporal pathology, whereas Korsakoff patients did not show disproportionate spatial memory impairment. Items located within an array were presented to Korsakoff and mixed aetiology amnesic patients, and to respective control groups. Recognition memory for the location of items was tested after incidental and intentional encoding. When equated on item recognition, the Korsakoff group did not benefit from intentional, relative to incidental, encoding instructions. Moreover, the Korsakoff patients did not show either disproportionately impaired incidental or intentional location memory, relative to their controls. By contrast, the mixed-aetiology patients profited significantly from intentional location learning, relative to incidental instructions, and were impaired on the incidental tasks, relative to normal control. It was argued that this group showed a disruption to the ability to bind item and location information.
Kopelman et al. (1997) compared temporal and spatial context memory in patients with Korsakoff pathology, temporal lobe, or focal frontal damage, identified with quantified MRI and FDG-PET neuroimaging. Patients were presented with two series of line drawings 45 min apart, and each line drawing was positioned at either the top or bottom of the slide. Subsequently, the participants performed a Yes-No item recognition test, and temporal and spatial judgement tasks. The groups were matched for the level of recognition memory performance, by providing prolonged exposure times to the memory-disordered patients, compared with healthy controls. Patients with Korsakoff’s syndrome showed significant impairments on the temporal context (recency) task, as did patients with frontal lesions affecting the lateral cortical margins. On a spatial context memory task, patients with lesions in the temporal lobes showed significant impairment, and patients with right temporal lesions performed significantly worse than patients with left temporal lobe lesions. There were strong correlations between the severity of anterograde memory impairment, determined on standard tasks, and context memory performance and only weak associations with executive tasks. In summary, this study was consistent with the dissociations postulated by Parkin and Chalfonte, although these dissociations were not entirely ‘clean’: examination of the raw data clearly suggested that the temporal lobe group showed impairment at the temporal context task, and the Korsakoff group at the spatial context task, although neither impairment reached statistical significance.
More Recent Investigations
In a study by Kessels et al. (2000), different aspects of object location memory were examined in a group of Korsakoff patients and age- and education-matched controls. They compared three task conditions: memory for pure locations (without any objects present); object-location binding (a cued-recall task condition in which objects had to be placed on their original positions that were pre-marked by dots); and a condition in which objects had to be located within an empty frame as precisely as possible (without any pre-marked locations). In addition, they examined recognition memory for items. Results demonstrated large deficits on all spatial-memory conditions, with no evidence for selectivity. Further analyses of covariance demonstrated that while item recognition memory was also impaired in the Korsakoff group, the deficits on the spatial memory conditions could not be fully explained by the general item-memory deficit. These results are in line with Mayes et al.’s (1985) context-deficit hypothesis, and Shoqeirat and Mayes’s (1991) findings.
A later study (Postma et al. 2006) directly compared memory for the location and temporal order of objects in a group of Korsakoff patients. Here, they also varied the presentation and recall conditions. Spatial presentation of objects (comparable to Kessels et al. 2000) was followed by a spatial recall phase, and temporal presentation of objects (at one location) was followed by a temporal-order recall condition. In addition, the spatiotemporal presentation conditions were either followed by a purely spatial or temporal recall phase, or a combined spatiotemporal recall phase. The results showed that the Korsakoff patients and the controls had comparable deficits for spatial and temporal-order recall in the single-feature conditions. More interestingly, a profound difference was found in the Korsakoff patients on the combined recall conditions. The authors argued that these results extended the context memory deficits hypothesis, in that memory for contextual processing was indeed impaired in Korsakoff’s syndrome, and that this deficit was enhanced where multiple features had to be bound, as in the combined recall conditions.
Most recently, Tielemans et al. (2012) have examined the facilitation of memory by contextual cues in patients with diencephalic or medial temporal pathology. They included twenty patients with Korsakoff’s syndrome, twenty patients with unilateral medial temporal lobectomy, and 36 healthy volunteers. Participants had to learn forty nouns, randomly combined with a photograph of a scene. In a cued recall task, the cue was presented alongside the same or a different scene. On a recognition memory test, each of the contextual scenes was presented in association with one of the target words, and one of the distractors. The authors found that, on the cued recall task, the contextual cues did not facilitate performance in the Korsakoff patients, suggesting an impairment in item-context binding during this task. The Korsakoffs did not show an impairment during the recognition memory test, and the unilateral temporal lobectomy patients showed normal facilitation by contextual cues in the cued recall tasks. This suggests that the unilateral temporal lobe patients were able to respond to contextual cues in a way that the Korsakoff patients, with presumed bilateral pathology in the thalamic-fornix-hippocampal circuitry, could not. It should be acknowledged, however, that in this study no patients with bilateral medial temporal-lobe pathology were examined.
Pitel et al. (2008) examined 14 Korsakoff patients, 40 non-Korsakoff alcoholics, and 55 healthy controls, matched for age and education, on an ecological contextual memory task. Participants learned unrelated word pairs presented at different times and places within a single day. At the end of the day, they performed a recognition memory test of the factual, temporal, and spatial information. Korsakoff patients performed significantly worse than the controls and alcoholics at the factual, temporal and spatial components of the task.
Summary
In brief, there is now abundant evidence that both temporal context memory and spatial context memory are disproportionately impaired in Korsakoff patients. However, the degree of impairment in these two types of context memory does vary from study to study, and it may well be the case that differing pathways and projections are involved in these impairments (compare Postma et al. 2006). Projections to and from the lateral frontal cortices may be particularly critical in producing the temporal context memory impairment, and projections to and from the hippocampi (especially on the right) may be particularly critical to the spatial context memory deficit. Context memory is an important component of recollection (i.e., the conscious retrieval of information from long-term memory), and, in recent years, debate as to whether a context memory deficit underlies Korsakoff patients’ memory impairment has become assimilated into the wider debate about the role of the ‘extended hippocampal’ circuitry (Aggleton and Brown 1999) in recollection and familiarity-based memory processes.
Contextual Retrieval in the Korsakoff Syndrome
Contextual Cues in Retrograde Amnesia and Autobiographical Memory
As mentioned above, Kopelman (1989) showed a deficit in the temporal dating of famous news events in Korsakoff patients, relative to healthy controls, with a U-shaped curve, such that there was pronounced primacy and recency effects in dating recalled news events.
Mayes et al. (1994) carried out a related investigation, in which the authors described two tests for assessing remote memory in amnesic patients. Mayes and his co-authors argued that it was impossible to evaluate competing hypotheses about temporally-graded retrograde amnesia (relative sparing of early memories) unless items were selected from at least two difficulty levels and from at two time-periods. Based on these principles, the authors designed a test of famous people and a test of famous public events. On these tasks, they found only a weak temporal gradient over a 20-year period, not significant in terms of the group by delay interaction effects. However, when the authors examined patients’ dating of faces/events, they obtained disproportionate impairment in their amnesic group, which contained some Korsakoff patients, with a statistically significant temporal gradient (relative sparing of early memories). The patients’ memory for temporal dating was shown to be more severely affected than was their memory for other types of question. The authors interpreted this finding in terms of the context memory deficit hypothesis, arguing that this would indeed predict a disproportionate deficit in the dating of famous names and events, relative to recognition memory ability, and also relative to memory for other types of (non-contextual) information about the famous person or event acquired at the same time.
Parkin et al. (1990b) examined this issue more directly. They investigated the effects of contextual cueing on the remote memory of Korsakoff patients in comparison with healthy subjects. The participants were required to name famous personalities who had become famous across five decades, under each of two experimental conditions—a ‘no-context’ condition in which there were minimal extraneous cues to the identification of the famous personalities, and a ‘context’ condition where clear extraneous cues were available. Korsakoff patients were impaired overall on this test, and they showed a marked temporal gradient. Both patients and controls showed benefits from contextual cueing, which was less pronounced in more recent decades. The authors concluded that a context memory deficit could not explain the impairment of the Korsakoff patients’ retrograde amnesia.
In recent years, this issue has been subsumed within the debate about recollection and familiarity-based memory impairment, as in investigations of anterograde amnesia. Various authors have argued that memory disordered patients, such as traumatic brain injury and Alzheimer patients, show disproportionate impairment in autobiographical memory on measures of recollection involving detailed contextual memory recall, and that this recollection deficit has been attributed to damage in the hippocampi or extended hippocampal circuitry (Piolino et al. 2003, 2007; Rosenbaum et al. 2005, 2008). However, this issue has not been examined systematically in Korsakoff patients.
Temporal Confusion and Confabulation
In discussing confabulation, Korsakoff (1889) himself emphasised the role of real memories, recalled out of temporal sequence, such that they were retrieved inappropriately out of their temporal context. He identified a confusion of “old recollections with present impressions” as the basis of many instances of confabulation. He gave the example of a patient who “in telling of something about the past, the patient would suddenly confuse events and would introduce events related to one period into the story about another period…Telling of a trip she had made to Finland before her illness and describing her voyage in fair details, the patient mixed into the story her recollections of Crimea, and so it turned out that in Finland, people always eat lamb and the inhabitants are Tartars.” Many other clinical studies adopted a similar interpretation of confabulation (Moll 1915; Victor et al. 1971; Talland 1965; Van Der Horst 1932).
Berlyne (1972) and Kopelman (1987) distinguished between momentary or provoked confabulation and spontaneous confabulation. The former referred to brief or fleeting intrusion errors or distortions when memory was challenged, and Kopelman (1987) argued that this was a normal property of ‘weak’ memory. Spontaneous confabulation referred to a persistent, provoked outpouring of erroneous memories. Kopelman (1987) argued that spontaneous confabulation was a pathological phenomenon, resulting from frontal pathology, and involving an extremely incoherent and context-free retrieval of memories and associations. In a subsequent detailed case report, Kopelman et al. (1997) examined a Korsakoff patient who also had a paraneoplastic syndrome, finding that this patient confabulated floridly across autobiographical episodic memory (events), personal semantic memory (facts), and orientation tasks. However, there appeared to be a number of underlying mechanisms, such that many (but not all) of the confabulations in autobiographical episodic memory appeared to reflect temporal context memory errors, whereas the confabulations in semantic memory often reflected perseverations.
Schnider et al. (1996) tested the temporal context confusion hypothesis much more directly (see also Schnider et al. 2000; Schnider 2003). These researchers demonstrated that spontaneous confabulators (defined in terms of their having acted upon their confabulations) could be differentiated from other amnesic patients and from healthy controls on the basis of their errors at a temporal context memory task, but not on other memory and executive tests. In their task (Schnider et al. 1996), there were two ‘runs’ of a continuous recognition memory test. In the second ‘run’, previous distractors became targets, and targets became distracters. This method provoked false positive responses in saying which items had been seen before within a given ‘run’. In Schnider et al.’s (1996) experiment, the confabulating patients were clearly differentiated from non-confabulating amnesic patients and healthy controls on this task. Moreover, this very specific deficit subsided as the confabulation improved. Subsequently, Treyer et al. (2003), Treyer, Buck and Schnider (2003), Schnider et al. (2007), and Schnider et al. (2005) employed PET-imaging and event-related potential (ERP) data to identify the malfunctioning of a very rapidly acting (200–300 milliseconds) filter, located within the orbitofrontal cortex, which, they argued, sets the ‘cortical format’ for subsequent memory encoding.
Gilboa et al. (2006) attempted a replication of this study. Using Schnider’s technique, they found that the temporal context memory task did indeed appear to differentiate confabulating memory-disordered patients from non-confabulating patients with medial temporal lobe pathology. Their group of confabulators had anterior communicating artery aneurysms. However, when these authors compared their confabulators’ performance with that of non-confabulating patients with comparable anterior communicating artery aneurysms, they found that the latter group’s performance on the Schnider test overlapped with that of the confabulating group, i.e. the test was not discriminating. Moreover, when the authors adapted the task so that participants had to make difficult discriminations (involving alternative views of objects) which did not involve a temporal context judgement, they identified a similar pattern—namely, the confabulators were differentiated from the medial temporal lobe pathology group, but non-confabulating patients with anterior communicating artery aneurysms showed performance which overlapped with both the confabulators and the medial temporal group. In recent study as yet unpublished, Bajo et al. (2012) have obtained a related finding in 24 confabulators, non-confabulating brain-injured memory-disordered patients, and healthy controls. In general, but not always, the Schnider test differentiated between confabulators and healthy controls: brain-injured non-confabulating amnesic patients showed performance on the Schnider test which overlapped with both the other two groups. In summary, temporal context confusion, as measured on Schnider’s test, may contribute to, but cannot alone account for spontaneous confabulation.
Summary
There is evidence that a context memory deficit (e.g. in dating events) is an important component of the retrograde amnesia in Korsakoff patients, and that it may contribute to the deficit in recollection, but the latter is not definitively established in this group of patients. More specifically, spontaneous confabulation has been attributed to a temporal context memory deficit, but this does not appear to be the full story.
Contextual Working Memory in Korsakoff’s Amnesia
Intact vs. Impaired Working Memory Function in Korsakoff’s Syndrome
In contrast to the severe deficits in long-term memory, both in antegrograde and retrograde memory, Korsakoff patients have long been assumed to perform normally on tasks relying on the temporary maintenance of information. For example, Baddeley and Warrington (1970) examined short-term memory in a group of amnesia patients (predominantly) due to Korsakoff’s syndrome. They used different measures of short-term memory, that is they examined the recency effect on a free-recall and paired-associate memory task, the rate of forgetting after an interference tasks (i.e., the Brown-Peterson paradigm in which participants have to count backwards during maintenance of a stimulus set) and performance on the digit span task, in comparison to a matched control group. On none of these measures did the Korsakoff patients demonstrate a worse performance than the controls. This finding has been frequently (but not always) replicated since (see Kopelman 1991b, 2002, for an overview) and has provided further evidence for the dissociation between short-term and long-term memory, both at a functional and a neurocognitive level. However, studies showing intact performance on working-memory tasks in Korsakoff patients typically use tasks that rely on the passive short-term retention of information. In Baddeley and Hitch’s (1974) model, these tasks rely on the passive slave systems, the phonological loop for auditory-verbal information and the visuospatial sketchpad for visual and/or spatial information, coordinated by a ‘central’ executive control system. Typical for these slave systems is that they encompass a processing unit specialized in a single modality (e.g., spatial or verbal information) and cannot hold representation of multimodel or item-context information. A later addition to the mode, the episodic buffer (Baddeley 2000), could handle these multidimensional types of information. Basically, it acts like a short-term buffer that holds integrated information from different slave systems (such as object and location information) as a new temporary “episodic representation”, yet being independent from the long-term memory system (see Baddeley 2000, 2012 for a more detailed discussion of the specific component, but see Gooding et al. 2005). Figure 2 shows the most recent version of this model (Baddeley 2012).

Baddeley’s working memory model (adapted from Baddeley 2012). The episodic buffer integrates information from the visuospatial sketchpad (VSSP) or phonological loop and holds it temporarily as a bound episodic representation, linking working memory to long-term memory (under the control of the central executive)
Several findings on working-memory performance in Korsakoff patients can be interpreted in the light of Baddeley’s revised model. First, Joyce and Robbins (1991) studied a group of Korsakoff patients on two computerized working-memory paradigms from the Cambridge Neuropsychological Test Automated battery (CANTAB). The first is the spatial span task, based on Corsi’s (1972) visuospatial analogue of the digit span, which assesses the capacity of the visuospatial sketchpad. The second is the “blue token” spatial-working memory task, in which participants had to search for a hidden (blue) token by clicking boxes that are presented on a computer screen. Clicking a box results in its opening; the box could then be empty or filled with a blue token. The box then closes again and the participant has to search for a new hidden token, with the restriction that previous target boxes remain filled and this cannot contain one of the new tokens. The number of to be searched boxes gradually increases and two types of errors can be made: “within-search” errors occur if a participants returns to a previously opened (empty) box within a search and “between-search” errors occur if a participants opens a box that already contained a target item in a previous search. In their study, Joyce and Robbins (1991) showed that the Korsakoff patients were unimpaired on the Corsi task (compare Kopelman 1991b), but showed profound deficits on the spatial-working memory paradigm. That is, especially in the high-load trials (6 or 8 boxes) did the Korsakoff patients perform increasingly worse than matched controls, but only on the between-search errors. The authors explain this deficit by an impaired search efficiency in Korsakoff patients, as reflected in a strategy measure (i.e., the consistency of the search path in each search) of the spatial working-memory paradigm as well as impaired performance on a planning task that relies on executive function.
The Role of the Episodic Buffer in Contextual Working Memory
However, one could also argue that the previous target locations must be kept in the episodic buffer, as the between-search errors reflect the inability to maintain target-location information over longer periods of time. To investigate this, a modified version of this working-memory task (the Box Task; Van Asselen et al. 2005) was developed, which enabled the presentation of different target items (everyday objects) instead of only one blue token. Using this paradigm, Korsakoff patients consistently produced more between-search errors in the high-load trials, in the absence of a within-search deficit or a deficit on the spatial span as measured with the Corsi Block-Tapping Test (Van Asselen et al. 2005; Oudman et al. 2011). Interestingly, Van Asselen et al. (2005) also examined the search strategy using a similar measure to that used by Joyce and Robbins (1991, based on Feigenbaum et al. 1996). On this measure, no difference was found between the Korsakoff patients and the matched controls. The role of context was further investigated by van Asselen et al. (2005) as two task conditions were administered: one with all boxes being identical and one in which all boxes had different colours. In the latter conditions, these colours could serve as contextual cues in that not only the location but also the colour provides information on already searched and previous target boxes. However, although the controls’ performance greatly improved in this contextual cueing condition, the Korsakoff patients did not benefit from these additional cues. In summary, Korsakoff patients were impaired on spatial working memory paradigms in which target-location information had to be maintained across trials, where it can be argued that performance relies on the binding function of the episodic buffer. In turn, no deficits were present in Korsakoff patients on tasks that relied on the maintenance of locations that required only processing at the level of the visuospatial sketchpad, (i.e. as reflected by the low number of within-search errors on the spatial working memory task or by intact performance on Corsi-like paradigms).
Whereas these studies provide evidence for a role of the episodic buffer, one might still argue that these are predominantly due to executive deficits, i.e. the inability to effectively search trough the presented boxes. However, deficits in Korsakoff patients have also been demonstrated on other working memory tasks. For example, Brokate et al. (2003) showed a deficit on the 2-back condition of the n-back task, a test that may rely on the episodic buffer’s ability to maintain the temporal position across trials (Kessels et al. 2010) and that has been shown to be independent from the working memory slave system (Kane et al. 2007). Furthermore, working memory for single versus multiple features has been examined using a Sternberg paradigm (Piekema et al. 2007). In that study, Koraskoff patients performed worse on all task conditions, both on the single-feature trials (colour or location only) and the multiple-feature (digit-colour or digit-location) associations. Possibly, this overall working memory deficit might have been the result of the long delay that was used (8 s), supported by the notion that almost all patients performed unimpaired on the Corsi Block-Tapping Task in this study. Moreover, Oscar-Berman and colleagues (Oscar-Berman et al. 1982, 1992) had previously shown that Korsakoff patients perform poorly on delayed response tasks with short delays (5–30 s) (but contrast Baddeley and Warrington 1970; Kopelman 1991b). The study by Piekema et al. (2007) was followed up with a systematic comparison of working memory for bound representations using a Sternberg paradigm (in which a set of face-house pairs had to be maintained, followed by a probe) both at a 3 s or a 6 s delay (Van Geldorp et al. 2012). Here, Korsakoff patients were also impaired compared to controls, but no effect of delay was found. However, in this study working memory for single features was not examined. Finally, Pitel et al. (2008) reported impaired performance in Korsakoff patients on a multimodal span task (object-location working memory) that was used to tap the episodic buffer. However, in this study, Korsakoff patients performed worse than healthy controls on slave system working memory tasks as well, which the authors related to the prefrontal damage in these patients.
Neural Correlates and Role of Long-Term Encoding
Thus, Korsakoff patients generally show intact performance on “context-free” working memory paradigms that rely on slave system processing (Baddeley and Hitch 1974), such as simple span tasks or tests that require immediate recall of single items. Presumably, working memory processing at the level of Baddeley’s (2000) slave systems relies at least in part on the prefrontal cortex. As a result, inconsistencies in the literature arise (e.g. impaired performance of Korsakoff patients on simple span tasks) and may be the result of differing degrees of prefrontal damage due to chronic alcohol abuse in Korsakoff patients independent of diencephalic-hippocampal dysfunction (Kopelman 1985, 1991a,b; Joyce and Robbins 1991; Pitel et al. 2011). Working-memory tasks that require contextual processing (such as working memory for object locations or the maintenance of temporal positions across trials) consistently produce large deficits in Korsakoff patients, which may be the result of dysfunction of the episodic buffer. The underlying neural correlate of this episodic buffer deficit could be the extended hippocampal system, which is crucial for the formation of new episodes in long-term memory and which may also be important for relational working memory at short delays (e.g., Piekema et al. 2009; Ranganath and Blumenfeld 2005; Olson et al. 2006). While the hippocampus may directly be affected in some cases of the Korsakoff syndrome (Sullivan and Pfefferbaum 2009), non-Korsakoff patients with selective bilateral hippocampal lesions have been shown to be unimpaired on relational working memory tasks (Baddeley et al. 2011). Possibly, only lesions in the wider dicencephalic-hippocampal circuit result in deficits in contextual working memory.
In turn, one could argue that the impairments on contextual working memory tasks merely reflect a deficit in long-term encoding (by definition present in Korsakoff amnesics). This view is supported by evidence that the extent of the working memory deficit increases with longer delays (Olson et al. 2006). However, Oscar-Berman et al. (1992) already demonstrated that the short-term delayed response deficit in Korsakoff patients was unrelated to the degree of long-term memory dysfunction (as measured by Wechsler’s MQ). In addition, Van Geldorp et al. (2012) have examined episodic memory formation of the stimuli presented in the relational Sternberg working memory task using a surprise subsequent memory test. Although the Korsakoff patients performed poorly on the subsequent memory test, performance was related neither to the level of working memory performance in patients or controls nor to the delay length. It could be argued that a delay of 6 s would enhance long-term encoding processing during working-memory maintenance in the healthy controls compared to the shorter (3 s) delay, thus resulting in a better subsequent memory performance. However, no delay effect was found in the control group as well.
Summary
In all, there is clear evidence that Korsakoff patients show working memory deficits for contextual information that extends beyond the slave systems, such as multi-dimensional, associative representations that must be kept online across trials over longer periods of time. These representations may be mediated by the episodic buffer (cf. Baddeley 2000, 2012). This deficit is likely to result from dicencephalic-hippocampal dysfunction typical of Korsakoff’s syndrome, as opposed to slave-system impairments which in Korsakoff patients may originate from prefrontal atrophy secondary to chronic alcohol abuse (also present in non-Korsakoff chronic alcoholics).
Implicit Contextual Memory
Intact Implicit Memory Processing
Early in the twentieth century, it was already recognized that not all aspects of memory processing were impaired in patients with Korsakoff’s amnesia. As early as 1907 (Claparède 1907) the Swiss psychologist Claparède demonstrated unconscious memory. In an experiment, he showed a female Korsakoff patient who was densely amnesic ten random words that she had to read aloud. After having read them, a free recall memory test followed. This was repeated seven times. The experiment was repeated with the same words on different days. The results demonstrated a clear learning ability: in order to learn to a criterion of 7 words, the first session required 5 readings, on the second day only two readings were necessary, and on the third only one reading. Preserved unconscious learning was later demonstrated by Claparède (1911) in his classic anecdote in which he pricked the same patient with a hidden pin in his hand. Later, she reflexively withdrew her hand when meeting him, without having a conscious recollection. A similar description was given by Korsakoff himself (1889), providing anecdotal evidence for “weak memories” that could influence behaviour outside consciousness. Later, observations like these and numerous experiments formed the basis for a dissociation between explicit, episodic memories for events and implicit, unconscious memory. These dissociations have sometimes been interpreted as discrete memory systems, and sometimes as different types of memory processing (see Schacter 1987, for an overview). Typically, all classic forms of implicit memory have been demonstrated to be spared in Korsakoff’s amnesia, such as perceptual learning, repetition priming, and procedural learning (Kopelman et al. 2009; d’Ydewalle and Van Damme 2007).
However, all these forms of implicit memory are context-free; that is, they do not rely on contextual processing, such as item-location processing or inter-item context learning. As the previous sections of this paper has demonstrated clear deficits in several forms of context memory, both in working memory and in episodic memory, the question arises whether implicit learning of contextual information is spared or not. For example, explicit object-location memory relies on the diencephalic-hippocampal memory system in man, but “automatic” forms of spatial learning in rodents also require an intact hippocampal circuit (Kesner and Goodrich-Hunsaker 2010). Moreover, different forms of contextual processing may facilitate later behaviour. For example, Verfaellie et al. (2008) did not find an enhanced repetition effect in Korsakoff patients for words that were repeatedly presented using a varied semantic context (different semantically related words for each presentation of the target) compared to words presented in the same semantic context at study (i.e., the same semantically related word for each target). Varied context presentation is assumed to provide multiple retrieval cues that facilitate later recollection. Whereas this was the case for healthy older adults, both conditions resulted in an increased feeling of knowing in Korsakoff patients to a similar extent.
Implicit Learning of Contextual Information: Word Pairs and Object Locations
In this section we will focus on the acquisition of information that in itself is contextual, such as relational information in the verbal (word pairs) or visuospatial (object-location associations) domain. Van Damme and d’Ydewalle (2008) studied implicit semantic contextual processing in Korsakoff patients by presenting words that were frequently (typical) and infrequently (atypical) paired with semantically related words. Using Jacoby’s (1991) process-dissociation procedure (PDP), the contribution of conscious recollection (R) and implicit processing (habit or H) were estimated. A large difference between Korsakoff patients and matched controls was found on the estimate of R, whereas the estimates of H were minimally different between the two groups. A second experiment in which Korsakoff patients were given more time to respond even reduced the large difference between the groups for R. These findings showed that Korsakoff patients were able to benefit from semantic contextual information.
The process dissociation procedure (PDP) was also applied in a study by Postma et al. (2008), who examined implicit and explicit processing for object locations in 23 Korsakoff patients. They presented everyday objects in specific locations in photographed rooms on a computer screen at study. During the test phase, specific instructions were given to the participants in that they had either to place an object at the location it previously occupied at study (“include trials”) or had to place them at a different location than the previously occupied one (“exclude trials”). If they could not remember the previously occupied location, participants were instructed to place them at the first location that came to mind. Using this procedure, estimates of conscious and implicit memory could be determined. Furthermore, all participants were re-tested after 1 week with the same instructions. Here, the results again demonstrated a large group difference in conscious memory, whereas both groups had an equal estimate of implicit memory. Interestingly, and as expected, after 1 week, the contribution of conscious recollection deteriorated substantially in both groups, whereas the contribution of implicit processing to the response remained stable in both groups. These findings provided further evidence that Korsakoff patients were able to process contextual information in an implicit manner, but have difficulty with strategic and conscious retrieval.
Spatial Motor Learning
The processing of spatial contextual information is crucial in many everyday motor tasks. That is, motor behaviour has a profound spatial component, as it often relies on movements that have to be made within a two-dimensional space (such as drawing or pressing buttons on an electrical device) or three-dimensional space (such as moving around in an actual environment). Several studies have examined the effects of spatial priming in Korsakoff patients using motor-learning paradigms. In 1992 (Verfaellie et al. 1992), Verfaellie, Milberg, Cermak and Letourneau studied the effect of priming of spatial configurations (interfering connected-dot patterns that had to be copied between study and test) on later recall involving drawing of previously studied connected-dot patterns. By comparing Korsakoff patients with alcoholic and non-alcoholic controls, this study demonstrated spatial priming of the copied interfering dot patterns on the final drawings in all three groups, albeit to a lesser extent in the Korsakoff patients (who as a result showed reduced interference).
Priming effects of spatial context in Korsakoff patients were also examined by Oudman et al. (2011). They showed participants subsequent spatial displays that contained one target item (a T) among 11 distracter items (Ls) presented in different colours in different orientations. The participant’s task was to locate the target item as soon as possible and determine the orientation (90 ° rotated or 270 °) by button press in every trial. Some of the spatial configurations were repeated during the blocks, and the difference in reaction time between repeated and novel displays was used as an estimate of implicit learning ability. In general, the Korsakoff patients were slower in responding than the matched controls, but the spatial priming effect was similar in both groups, suggesting that Korsakoff were patients are able to use learn contextual information. This study also extended previous research in which the same task was administered in patients with damage to the medial temporal lobe including the hippocampus (Chun and Phelps 1999) and patients with selective damage to the hippocampal formation (Manns and Squire 2001). The medial temporal-lobe patients did not show implicit contextual learning on this task, whereas the patients with lesions limited to the hippocampal formation showed preserved contextual learning (similar to the Korsakoff patients). It is likely that other medial temporal lobe structures, such as the entorhinal, perirhinal and parahippocampal cortex, may be more important for implicit contextual processing than the hippocampus proper or dicenephalic structures such as the mammillary bodies or the thalamus. Van Tilborg et al. (2011) have also shown intact implicit motor learning in Korsakoff patients on both a spatially complex motor learning task (pattern learning) and a classic serial reaction time task with a less complex spatial layout.
Although several studies have been performed on implicit learning of computerized spatial motor tasks in Korsakoff patients, surprisingly little research has been performed on actual route learning. Geilenkirchen (1990) examined actual route learning in ten Korsakoff patients, in which patients walked the same route together with the researcher on subsequent days (5 trials in a 3-week period). This study showed largely preserved route learning in most patients, although only three patients managed to acquire the route completely. Later Kessels et al. (2007) also investigated route learning in Korsakoff patients in a study in which an actual route was walked under two learning conditions on multiple days (errorless vs. trial-and-error learning). No difference between the two learning conditions was found, and clear evidence of learning was found in all patients (although it must be noted that not all patients acquired the complete route).
Summary
While there is abundant evidence for intact implicit learning in patients with amnesia due to Korsakoff’s syndrome, less attention has been given to implicit contextual learning. Studies on incidental context cues present during learning and/or retrieval have demonstrated that Korsakoff patients benefit from such contextual cues only if these are distinctive (such as a dimly lit room with background music playing). Implicit learning of information which in itself is contextual (for instance, word pairs or associations between objects and their locations) is well-preserved in Korsakoff patients. In addition, route learning in actual environments that rely on implicit (procedural) contextual information processing is also relatively intact in Korsakoff patients, although this has been scarcely studied only in a very few investigations to date.
Conclusion
This review clearly has demonstrated that patients with Korsakoff’s syndrome have deficits in the explicit processing of contextual information. Specifically, the ability to store contextual information (such as spatial locations or temporal order information) and to use contextual information as a retrieval cue is impaired. Moreover, an additional deficit may exist in the binding of contextual information to targets. These findings largely corroborate and extend Mayes et al.’s (1985) context deficit hypothesis, which argued that the dicencephalic-hippocampal memory system is crucial for contextual memory formation and retrieval. Results with respect to differences between different types of contextual information are somewhat mixed, but there is evidence showing that Korsakoff patients have difficulties with both memory for temporal order and for spatial memory. Context memory impairments may also contribute to the loss of ‘recollective’ remote memories, and their dating, and to the production of confabulations in the episodic memory (Schnider et al. 1996; Kopelman et al. 1997), although other factors such as defective monitoring (Gilboa et al. 2006) and affective motivation (Fotopoulou et al. 2008) may also contribute to confabulation.
Furthermore, we have clearly demonstrated deficits in contextual working memory, that is the ability to maintain associative information over a brief period of time. It is still under debate whether this associative working memory deficit in Korsakoff patients simply reflects an impairment of long-term encoding processes taking place within working-memory tasks (and perhaps reflecting the activity of the extended hippocampal system) or whether this is the result of a “pure” working memory deficit (for instance, dysfunction of the episodic buffer).
In contrast to the profound contextual deficits in conscious, declarative forms of memory (working memory and episodic memory) in Korakoff’s syndrome, implicit, nondeclarative contextual learning is largely preserved in these patients. Korskaoff patients benefit from implicit contextual cues during learning, and are able to acquire complex spatial motor patterns. Moreover, there is some evidence that they are able to learn a route in actual environments. Thus, a dissociation seems to be present between impaired explicit contextual memory and intact implicit contextual learning. These findings are in line with the notion that explicit and implicit memory function (whether regarded as ‘systems’ or ‘processes’) rely on separate neural circuits, that is, the diencephalic-hippocampal memory system for explicit memory, and subcortical structures such as the basal ganglia for implicit, procedural learning (Squire and Zola 1996; Aggleton and Brown 1999; Wang and Morris 2010). In turn, it has also been argued that this dissociation is less related to the level of consciousness involved, but that many implicit tasks rely more on perceptual processing than elaborative semantic processing (Brunfaut and d’Ydewalle 1996). However, in the case of Korsakoff’s syndrome, explicit contextual memory processing for visuospatial information is clearly impaired, suggesting that indeed implicit contextual processing is mediated by a different neural circuit.
One of the problems with unravelling the underlying neurocognitive mechanisms of contextual memory in Korsakoff’s syndrome is that most of the studies so far have adopted a primarily behavioural approach, and there have been few functional imaging investigations. Lesions in the diencephalon have been consistently shown in post-mortem research studies (Mair et al. 1979; Mayes et al. 1988; Harding et al. 2000). In structural MRI investigations, atrophy of the thalami, mammillary bodies, and prefrontal cortex have been found (Colchester et al. 2001; Sullivan and Pfefferbaum 2009) and of the hippocampi in one of these studies (Sullivan and Marsh 2003) but not the other (Colchester et al. 2001). On FDG-PET, there is reduced glucose uptake in the thalamus, hypothalamus, basal forebrain, and retrosplenium (Reed et al. 2003). As a result, it is difficult to disentangle the role of prefrontal dysfunction and diencephalic-hippocampal dysfunction in the memory deficits in Korsakoff patients. Notably, the deficits in contextual working memory have been linked to prefrontal dysfunction, although deficits in executive function were in most studies unrelated to the working-memory deficits in Korsakoff patients. With respect to the anterograde amnesia, there is little doubt that these are due to dysfunction of the diencephalic-hippocampal memory system, and the retrograde amnesia probably involves dysfunction in the frontal lobes as well (Kopelman 1991a; Kopelman et al. 2003). With respect to the underlying neural mechanisms of implicit contextual learning, a dissociation exists between patients with selective hippocampal lesions and Korsakoff patients on the one hand (showing preserved contextual learning), and medial temporal-lobe patients on the other hand (who show impaired contextual learning). This may indicate the importance of non-hippocampal medial temporal-lobe structures that are largely preserved in Korsakoff’s syndrome in implicit contextual learning.
References
Aggleton, J. P., & Brown, M. W. (1999). Episodic memory, amnesia, and the hippocampal-anterior thalamic axis. The Behavioral and Brain Sciences, 22, 425–444.
Baddeley, A., Jarrold, C., & Vargha-Khadem, F. (2011). Working memory and the hippocampus. Journal of Cognitive Neuroscience, 23, 3855–3861.
Baddeley, A. D. (2000). The episodic buffer: a new component of working memory? Trends in Cognitive Science, 4, 417–423.
Baddeley, A. (2012). Working memory: theories, models, and controversies. Annual Review of Psychology, 63, 1–29.
Baddeley, A. D., & Hitch, G. J. (1974). Working memory. In G. H. Bower (Ed.), The psychology of learning and motivation (Vol. 8). London: Academic.
Baddeley, A. D., & Warrington, E. K. (1970). Amnesia and the distinction between long- and short-term memory. Journal of Verbal Learning and Verbal Behavior, 9, 176–189.
Bajo, A., Fleminger, S., & Kopelman, M. D. (2012). Longitudinal studies of confabulation. Manuscript in preparation.
Berlyne, N. (1972). Confabulation. The British Journal of Psychiatry, 120, 31–39.
Brokate, B., Hildebrandt, H., Eling, P., Fichtner, H., Runge, K., & Timm, C. (2003). Frontal lobe dysfunctions in Korsakoff’s syndrome and chronic alcoholism: continuity or discontinuity? Neuropsychology, 17, 420–428.
Brunfaut, E., & d’Ydewalle, G. A. (1996). comparison of implicit memory tasks in Korsakoff and alcoholic patients. Neuropsychologia, 34, 1143–1150.
Butters, N., & Cermak, L. S. (1980). Alcoholic Korsakoff’s syndrome: An information-processing approach to amnesia. New York: Academic.
Chalfonte, B. L., Verfaellie, M., Johnson, M. K., & Reiss, L. (1996). Spatial location memory in amnesia: binding item and location information under incidental and intentional encoding conditions. Memory, 4, 591–614.
Chun, M. M., & Jiang, Y. (2003). Implicit, long-term spatial contextual memory. Journal of Experimental Psychology: Learning, Memory, and Cognition, 29, 224–234.
Chun, M. M., & Phelps, E. A. (1999). Memory deficits for implicit contextual information in amnesic subjects with hippocampal damage. Nature Neuroscience, 2, 844–847.
Claparède, E. (1907). Expériences sur la mémoire dans un cas de psychose de Korsakoff. Revue Médicale de la Suisse Romande, 27, 301–303.
Claparède, E. (1911). Recognition et moïté. Archives de Psychologie, 11, 79–90.
Colchester, A., Kingsley, D., Lasserson, D., Kendall, B., Bello, F., Rush, C., Stevens, T. G., Goodman, G., Heilpern, G., Stanhope, N., & Kopelman, M. D. (2001). Structural MRI volumetric analysis in patients with organic amnesia, 1: methods and comparative findings across diagnostic groups. Journal of Neurology, Neurosurgery, and Psychiatry, 71, 13–22.
Corsi, P. M. (1972). Human memory and the medial temporal region of the brain. Dissertation Abstracts International, 34(02), 891B.
d’Ydewalle, G., & Van Damme, I. (2007). Memory and the Korsakoff syndrome: not remembering what is remembered. Neuropsychologia, 45, 905–920.
Feigenbaum, J. D., Polkey, C. E., & Morris, R. G. (1996). Deficits in spatial working memory after unilateral temporal lobectomy in man. Neuropsychologia, 34, 163–176.
Fotopoulou, A., Conway, M. A., Solms, M., Tyrer, S., & Kopelman, M. D. (2008). Self-serving confabulation in prose recall. Neuropsychologia, 46, 1429–1441.
Geilenkirchen, M. (1990). Een onderzoek naar het effect van hulpmiddelen bij ruimtelijke oriëntatieproblemen van Korsakow-patiënten [A study on the effect of external aids on spatial orientation deficits in Korsakoff patients]. Unpublished Master Thesis, Rijksuniversiteit Limburg, Maastricht, The Netherlands.
Gilboa, A., Alain, C., Stuss, D. T., Melo, B., Miller, S., & Moscovitch, M. (2006). Mechanisms of spontaneous confabulations: a strategic retrieval account. Brain, 129, 1399–1414.
Gooding, P. A., Isaac, C. L., & Mayes, A. R. (2005). Prose recall and amnesia: more implications for the episodic buffer. Neuropsychologia, 43, 583–587.
Harding, A., Halliday, G., Caine, D., & Kril, J. (2000). Degeneration of anterior thalamic nuclei differentiates alcoholics with amnesia. Brain, 123, 141–154.
Hunkin, N. M., Parkin, A. J., & Longmore, B. E. (1994). Aetiological variation in the amnesic syndrome: comparisons using the list discrimination task. Neuropsychologia, 32, 819–825.
Huppert, F. A., & Piercy, M. (1976). Recognition memory in amnesic patients: effect of temporal context and familiarity of material. Cortex, 12, 3–20.
Huppert, F. A., & Piercy, M. (1977). Recognition memory in amnesic patients: a defect of acquisition? Neuropsychologia, 15, 643–652.
Huppert, F. A., & Piercy, M. (1978). The role of trace strength in recency and frequency judgements by amnesic and control subjects. The Quarterly Journal of Experimental Psychology, 30, 347–354.
Jacoby, L. L. (1991). A process-dissociation framework: separating automatic from intentional uses of memory. Journal of Memory and Language, 30, 513–541.
Joyce, E. M., & Robbins, T. W. (1991). Frontal lobe function in Korsakoff and non-Korsakoff alcoholics: planning and spatial working memory. Neuropsychologia, 29, 709–723.
Kane, M. J., Conway, A. R., Miura, T. K., & Colflesh, G. J. (2007). Working memory, attention control, and the N-back task: a question of construct validity. Journal of Experimental Psychology: Learning, Memory, and Cognition, 33, 615–622.
Kesner, R. P., & Goodrich-Hunsaker, N. J. (2010). Developing an animal model of human amnesia: the role of the hippocampus. Neuropsychologia, 48, 2290–2302.
Kessels, R. P. C., Meulenbroek, O., Fernández, G., & Olde Rikkert, M. G. M. (2010). Spatial working memory in aging and mild cognitive impairment: effects of task load and contextual cueing. Aging, Neuropsychology, and Cognition, 17, 556–574.
Kessels, R. P. C., Postma, A., Wester, A. J., & De Haan, E. H. F. (2000). Memory for object locations in Korsakoff’s amnesia. Cortex, 36, 47–57.
Kessels, R. P. C., Van Loon, E., & Wester, A. J. (2007). Route learning in amnesia: a comparison of trial-and-error and errorless learning in patients with the Korsakoff syndrome. Clinical Rehabilitation, 21, 905–911.
Kopelman, M. D. (1985). Rates of forgetting in Alzheimer-type dementia and Korsakoff’s syndrome. Neuropsychologia, 23, 623–638.
Kopelman, M. D. (1987). Two types of confabulation. Journal of Neurology, Neurosurgery, and Psychiatry, 50, 1482–1487.
Kopelman, M. D. (1989). Remote and autobiographical memory, temporal context memory and frontal atrophy in Korsakoff and Alzheimer patients. Neuropsychologia, 27, 437–460.
Kopelman, M. D. (1991a). Frontal dysfunction and memory deficits in the alcoholic Korsakoff syndrome and Alzheimer-type dementia. Brain, 114, 117–137.
Kopelman, M. D. (1991b). Non-verbal, short-term forgetting in the alcoholic Korsakoff syndrome and Alzheimer-type dementia. Neuropsychologia, 29, 737–747.
Kopelman, M. D. (2002). Disorders of memory. Brain, 125, 2152–2190.
Kopelman, M. D., Stanhope, N., & Kingsley, D. (1997). Temporal and spatial context memory in patients with focal frontal, temporal lobe, and diencephalic lesions. Neuropsychologia, 35, 1533–1545.
Kopelman, M. D., Lasserson, D., Kingsley, D. R., Bello, F., Rush, C., Stanhope, N., Stevens, T. G., Goodman, G., Buckman, J. R., Heilpern, G., Kendall, B. E., & Colchester, A. C. F. (2003). Retrograde amnesia and the volume of critical brain structures. Hippocampus, 13, 879–891.
Kopelman, M. D., Thomson, A. D., Guerrini, I., & Marshall, E. J. (2009). The Korsakoff syndrome: clinical aspects, psychology and treatment. Alcohol and Alcoholism, 44, 148–154.
Korsakoff, S. S. (1889). Etude médico-psychologique sur une forme des maladies de la mémoire. Révue Philosophique, 28, 501–530.
Mair, W. G., Warrington, E. K., & Weiskrantz, L. (1979). Memory disorder in Korsakoff’s psychosis: a neuropathological and neuropsychological investigation of two cases. Brain, 102, 749–783.
Manns, J. R., & Squire, L. R. (2001). Perceptual learning, awareness, and the hippocampus. Hippocampus, 11, 776–782.
Mayes, A. R. (1988). Human organic memory disorders. Cambridge: Cambridge University Press.
Mayes, A. R., & Downes, J. (1997). Theories of organic amnesia. Memory, 5, 1–2.
Mayes, A. R., Downes, J. J., McDonald, C., Poole, V., Rooke, S., Sagar, H. J., & Meudell, P. R. (1994). Two tests for assessing remote public knowledge: a tool for assessing retrograde amnesia. Memory, 2, 183–210.
Mayes, A. R., Meudell, P. R., & Pickering, A. (1985). Is organic amnesia caused by a selective deficit in remembering contextual information? Cortex, 21, 167–202.
Mayes, A. R., Meudell, P. R., Mann, D., & Pickering, A. (1988). Location of lesions in Korsakoff’s syndrome: neuropsychological and neuropathological data on two patients. Cortex, 24, 367–388.
Meudell, P. R., & Mayes, A. R. (1982). Normal and abnormal forgetting: some comments on the human amnesic syndrome. In A. W. Ellis (Ed.), Normality and pathology in cognitive functions (pp. 203–238). London: Academic.
Meudell, P., Mayes, A. R., Ostergaard, A., & Pickering, A. (1985). Recency and frequency judgements in alcoholic amnesics and normal people with poor memory. Cortex, 21, 487–511.
Moll, J. M. (1915). The amnesic or Korsakoff’s syndrome with alcoholic aetiology: an analysis of 30 cases. The Journal of Mental Science, 61, 423–437.
Montaldi, D., & Mayes, A. R. (2010). The role of recollection and familiarity in the functional differentiation of the medial temporal lobes. Hippocampus, 20, 1291–1314.
Nadel, L., & Moscovitch, M. (1997). Memory consolidation, retrograde amnesia and the hippocampal complex. Current Opinion in Neurobiology, 7, 217–227.
Olson, I. R., Page, K., Moore, K. S., Chatterjee, A., & Verfaellie, M. (2006). Working memory for conjunctions relies on the medial temporal lobe. The Journal of Neuroscience, 26, 4596–4601.
Oscar-Berman, M., Hutner, N., & Bonner, R. T. (1992). Visual and auditory spatial and nonspatial delayed-response performance by Korsakoff and non-Korsakoff alcoholic and aging individuals. Behavioral Neuroscience, 106, 613–622.
Oscar-Berman, M., Zola-Morgan, S. M., Oberg, R. G., & Bonner, R. T. (1982). Comparative neuropsychology and Korsakoff’s syndrome III: delayed response, delayed alternation and DRL performance. Neuropsychologia, 20, 187–202.
Oudman, E., Van der Stigchel, S., Wester, A. J, Kessels, R. P. C., & Postma, A. (2011). Intact memory for implicit contextual information in Korsakoff’s amnesia. Neuropsychologia, 49, 2848–2855.
Parkin, A. J., Leng, N. R. C., & Hunkin, N. M. (1990). Differential sensitivity to context in diencephalic and temporal lobe amnesia. Cortex, 26, 373–380.
Parkin, A. J., Montaldi, D., Leng, N. R., & Hunkin, N. M. (1990). Contextual cueing effects in the remote memory of alcoholic Korsakoff patients and normal subjects. The Quarterly Journal of Experimental Psychology, 42A, 585–596.
Piekema, C., Fernández, G., Postma, A., Hendriks, M. P. H., Wester, A. J., & Kessels, R. P. C. (2007). Spatial and non-spatial contextual working memory in patients with diencephalic or hippocampal dysfunction. Brain Research, 1172, 103–109.
Piekema, C., Kessels, R. P. C., Rijpkema, M., & Fernández, G. (2009). The hippocampus supports encoding of between-domain associations within working memory. Learning and Memory, 16, 231–234.
Piekema, C., Rijpkema, M., Fernández, G., & Kessels, R. P. C. (2010). Dissociating the neural correlates of intra-item and inter-item working-memory binding. PLoS One, 5, e10214.
Piolino, P., Desgranges, B., Belliard, S., Matuszewski, V., Lalevée, C., De La Sayette, V. D., & Eustache, F. (2003). Autobiographical memory and autonoetic consciousness: triple dissociation in neurodegenerative diseases. Brain, 126, 2203–2219.
Piolino, P., Desgranges, B., Manning, L., North, P., Jokic, C., & Eustache, F. (2007). Autobiographical memory, the sense of recollection and executive functions after severe traumatic brain injury. Cortex, 43, 176–195.
Pitel, A. L., Beaunieux, H., Witkowski, T., Vabret, F., de la Sayette, V., Viader, F., Desgranges, B., & Eustache, F. (2008). Episodic and working memory deficits in alcoholic Korsakoff patients: the continuity theory revisited. Alcohol Clinical and Experimental Research, 32, 1229–1241.
Pitel, A. L., Zahr, N. M., Jackson, K., Sassoon, S. A., Rosenbloom, M. J., Pfefferbaum, A., & Sullivan, E. V. (2011). Signs of preclinical Wernicke’s encephalopathy and thiamine levels as predictors of neuropsychological deficits in alcoholism without Korsakoff’s syndrome. Neuropsychopharmacology, 36, 580–538.
Postma, A., Antonides, R., Wester, A. J., & Kessels, R. P. C. (2008). Spared unconscious influences of spatial memory in diencephalic amnesia. Experimental Brain Research, 190, 125–133.
Postma, A., Van Asselen, M., Keuper, O., Wester, A. J., & Kessels, R. P. C. (2006). Spatial and temporal order memory in Korsakoff patients. Journal of the International Neuropsychological Society, 12, 327–336.
Ranganath, C., & Blumenfeld, R. S. (2005). Doubts about double dissociations between short- and long-term memory. Trends in Cognitive Science, 9, 374–380.
Reed, L. J., Lasserson, D., Marsden, P., Stanhope, N., Stevens, T., Bello, F., Kingsley, D., Colchester, A., & Kopelman, M. D. (2003). 18FDG-PET findings in the Wernicke-Korsakoff syndrome. Cortex, 39, 1027–1045.
Rosenbaum, R. S., Köhler, S., Schacter, D. L., Moscovitch, M., Westmacott, R., Black, S. E., Gao, F., & Tulving, E. (2005). The case of K.C.: contributions of a memory-impaired person to memory theory. Neuropsychologia, 43, 989–1021.
Rosenbaum, R. S., Moscovitch, M., Foster, J. K., Schnyer, D. M., Gao, F., Kovacevic, N., Verfaellie, M., Black, S. E., & Levine, B. (2008). Patterns of autobiographical memory loss in medial-temporal lobe amnesic patients. Journal of Cognitive Neuroscience, 20, 1490–1506.
Rudy, J. W. (2009). Context representations, context functions, and the parahippocampal-hippocampal system. Learning and Memory, 16, 573–585.
Schacter, D. L. (1987). Implicit expressions of memory in organic amnesia: learning of new facts and associations. Human Neurobiology, 6, 107–118.
Schnider, A. (2003). Spontaneous confabulation and the adaptation of thought to ongoing reality. Nature Reviews Neuroscience, 4, 662–671.
Schnider, A., von Däniken, C., & Gutbrod, K. (1996). The mechanisms of spontaneous and provoked confabulations. Brain, 119, 1365–1375.
Schnider, A., Mohr, C., Morand, S., & Michel, C. M. (2007). Early cortical response to behaviorally relevant absence of anticipated outcomes: a human event-related potential study. NeuroImage, 35, 1348–1355.
Schnider, A., Ptak, R., von Däniken, C., & Remonda, L. (2000). Recovery from spontaneous confabulations parallels recovery of temporal confusion in memory. Neurology, 55, 74–83.
Schnider, A., Treyer, V., & Buck, A. (2005). The human orbitofrontal cortex monitors outcomes even when no reward is at stake. Neuropsychologia, 43, 316–323.
Shoqeirat, M. A., & Mayes, A. R. (1991). Disproportionate incidental spatial-memory and recall deficits in amnesia. Neuropsychologia, 29, 749–769.
Smith, M. L., & Milner, B. (1981). The role of the right hippocampus in the recall of spatial location. Neuropsychologia, 19, 781–793.
Squire, L. R., Stark, C. E., & Clark, R. E. (2004). The medial temporal lobe. Annual Review of Neuroscience, 27, 279–306.
Squire, L. R., & Zola, S. M. (1996). Structure and function of declarative and nondeclarative memory systems. Proceedings of the National Academy of Sciences of the United States of America, 93, 13515–13522.
Sullivan, E. V., & Marsh, L. (2003). Hippocampal volume deficits in alcoholic Korsakoff’s syndrome. Neurology, 61, 1716–1719.
Sullivan, E. V., & Pfefferbaum, A. (2009). Neuroimaging of the Wernicke-Korsakoff syndrome. Alcohol and Alcoholism, 44, 155–165.
Talland, G. A. (1965). Deranged memory. New York: Academic.
Tielemans, N. S., Hendriks, M. P. H., Talamini, L., Wester, A. J., Meeter, M., & Kessels, R. P. C. (2012). Facilitation of memory by contextual cues in patients with diencephalic or medial temporal lobe dysfunction. Neuropsychologia.
Treyer, V., Buck, A., & Schnider, A. (2003). Subcortical loop activation during selection of currently relevant memories. Journal of Cognitive Neuroscience, 15, 610–618.
Tulving, E. (1983). Elements of episodic memory. Oxford: Clarendon.
Van Asselen, M., Kessels, R. P. C., Wester, A., & Postma, A. (2005). Spatial working memory and contextual cueing in patients with Korsakoff amnesia. Journal of Clinical and Experimental Neuropsychology, 27, 645–655.
Van Damme, I., & d’Ydewalle, G. (2008). Elaborative processing in the Korsakoff syndrome: context versus habit. Brain and Cognition, 67, 212–224.
Van Der Horst, L. (1932). Über die Psychologie des Korsakowsyndroms. Monatsschrift für Psychiatrie und Neurologie, 83, 65–84.
Van Geldorp, B., Bergmann, H. C., Robertson, J., Wester, A. J., & Kessels, R. P. C. (2012). The interaction of working memory performance and episodic memory formation in patients with Korsakoff’s amnesia. Brain Research, 1433, 98–103.
Van Tilborg, I. A. D. A., Kessels, R. P. C., Kruijt, P., Wester, A. J., & Hulstijn, W. (2011). Spatial and nonspatial implicit motor learning in Korsakoff’s amnesia: evidence for selective deficits. Experimental Brain Research, 214, 427–435.
Verfaellie, M., Milberg, W. P., Cermak, L. S., & Letourneau, L. L. (1992). Priming of spatial configurations in alcoholic Korsakoff’s amnesia. Brain and Cognition, 18, 34–45.
Verfaellie, M., Rajaram, S., Fossum, K., & Williams, L. (2008). Not all repetition is alike: different benefits of repetition in amnesia and normal memory. Journal of the International Neuropsychological Society, 14, 365–372.
Victor, M., Adams, R. D., & Collins, G. H. (1971). The Wernicke-Korsakoff syndrome. Oxford: Blackwell.
Visser, P. J., Krabbendam, L., Verhey, F. R., Hofman, P. A., Verhoeven, W. M., Tuinier, S., Wester, A., Den Berg, Y. W., Goessens, L. F., Van der Werf, Y. D., & Jolles, J. (1999). Brain correlates of memory dysfunction in alcoholic Korsakoff’s syndrome. Journal of Neurology, Neurosurgery, and Psychiatry, 67, 774–778.
Wang, S.-H., & Morris, R. G. M. (2010). Hippocampal-neocortical interactions in memory formation, consolidation, and reconsolidation. Annual Review of Psychology, 61, 49–79.
Warrington, E. K., & Weiskrantz, L. (1970). Amnesic syndrome: consolidation or retrieval? Nature, 228, 628–630.
Winocur, G., & Kinsbourne, M. (1978). Contextual cueing as an aid to Korsakoff amnesics. Neuropsychologia, 16, 671–682.
Witt, J. K., & Willingham, D. T. (2006). Evidence for separate representations for action and location in implicit motor sequencing. Psychonomic Bulletin Review, 13, 902–907.
Acknowledgments
Prof Kessels was supported by a VIDI grant from the Netherlands Organisation for Scientific Research (NWO, no. 452-008-005). Prof Kopelman’s work was supported by various grants from the Wellcome Trust.
Author information
Authors and Affiliations
Corresponding authors
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 2.0 International License ( https://creativecommons.org/licenses/by/2.0 ), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Kessels, R.P.C., Kopelman, M.D. Context Memory in Korsakoff’s Syndrome. Neuropsychol Rev 22, 117–131 (2012). https://doi.org/10.1007/s11065-012-9202-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11065-012-9202-5