
Abstract
Neurodevelopmental disorders such as attention deficit/hyperactivity disorder (ADHD) and autism spectrum disorder (ASD) are highly heritable and influenced by many single nucleotide polymorphisms (SNPs). SNPs can be used to calculate individual polygenic risk scores (PRS) for a disorder. We aim to explore the association between the PRS for ADHD, ASD and for Schizophrenia (SCZ), and ADHD and ASD diagnoses in a clinical child and adolescent population. Based on the most recent genome wide association studies of ADHD, ASD and SCZ, PRS of each disorder were calculated for individuals of a clinical child and adolescent target sample (N = 688) and for adult controls (N = 943). We tested with logistic regression analyses for an association with (1) a single diagnosis of ADHD (N = 280), (2) a single diagnosis of ASD (N = 295), and (3) combining the two diagnoses, thus subjects with either ASD, ADHD or both (N = 688). Our results showed a significant association of the ADHD PRS with ADHD status (OR 1.6, P = 1.39 × 10−07) and with the combined ADHD/ASD status (OR 1.36, P = 1.211 × 10−05), but not with ASD status (OR 1.14, P = 1). No associations for the ASD and SCZ PRS were observed. In sum, the PRS of ADHD is significantly associated with the combined ADHD/ASD status. Yet, this association is primarily driven by ADHD status, suggesting disorder specific genetic effects of the ADHD PRS.
Similar content being viewed by others


Introduction
Psychiatric disorders are heritable complex traits with varying heritability estimates. At the top end of the heritability range, reported heritabilities vary from 74% for ASD (Tick et al. 2016) to 80% for ADHD (Brikell et al. 2015), and 81% for SCZ (Sullivan et al. 2003). These traits likely have a similar genetic architecture with a role for common and rare variants, including de novo mutations and copy number variants (CNV) playing an important role (Gratten et al. 2014). Common genetic variation can be captured in a polygenic signal that contains a multitude of single nucleotide polymorphisms (SNPs) from many genes (Gratten et al. 2014; Sullivan et al. 2012). Genome-wide association studies (GWAS) are a highly successful method to identify the common variants that influence these disorders (Visscher et al. 2017). GWAS reveal increasingly more significantly associated loci. These represent the most associated part of the genetic signal. The most recent GWAS for ADHD, ASD, and SCZ identified 12, 5, and 145 independent associated loci, respectively (Demontis and Walters 2017; Grove et al. 2017; Pardiñas et al. 2018).
However, given the polygenicity of disorders like ADHD and ASD, also non-significantly associated SNPs are likely to contribute to the disorder (Wray et al. 2014). Hence, it is also of interest to investigate the non-genome-wide significant component of the genetic signal.
One method to include the non-genome-wide significant component of the common genetic variation is the polygenic risk scores (PRS) approach. PRS are the sum of risk alleles weighted by their estimated effect size as determined in an independent GWAS sample, and can serve as such as an estimation of an individual’s polygenic risk (Torkamani et al. 2018; Weiner et al. 2017; Wray et al. 2014). PRS estimated from an independent sample can be used for prediction between groups (e.g., cases and controls), or for stratifying groups of people according to high or low genetic risk as defined by their PRS. For example, in a sample of children from the general population, the SCZ PRS has shown positive associations with behavioral and emotional problems in children as young as 3 years old (Jansen et al. 2017). Similarly, the ADHD PRS has been associated with attention problems in children from the general population (Groen-Blokhuis et al. 2014), and with attentional and hyperactive-impulsive traits in another general population sample (age ~ 7 year, 7 months) (Martin et al. 2014).
As previous research indicates, the common genetic burden of different psychiatric disorders partially overlaps (Mitchell 2011). To add, both ADHD and ASD, as well as SCZ, are regarded neurodevelopmental disorders (NDD) (Mullin et al. 2013; Rapoport et al. 2012) and genetic studies have shown positive genetic correlations of 0.36 for ASD/ADHD (Grove et al. 2017), 0.211 for ASD/SCZ (Grove et al. 2017), and 0.122 for ADHD/SCZ (Demontis and Walters 2017).
In addition, it was shown that the prevalence of SCZ is significantly higher in an ASD sample compared to controls (OR 3.55, 95% CI 2.08–6.05, P < 0.001), and the prevalence of ASD in an SCZ samples ranges between 3.4 and 52% compared to 1% in the general population (Zheng et al. 2018). To add, ASD and SCZ share clinical features among which social cognition (Cheung et al. 2010; DSM 52013), while ASD and ADHD share inattention (Craig et al. 2015; DSM 52013).
The current study adds to this literature by investigating associations of the ADHD, ASD, and SCZ PRS in a sample of children and adolescents referred to an outpatient university clinic. The children in this sample were assessed with standardized procedures generating clinical (DSM-IV) diagnoses as well as continuous rating scale scores on behavioral/emotional problems. We aim to investigate whether PRS for ADHD (Demontis and Walters 2017), ASD (Grove et al. 2017) and SCZ (Pardiñas et al. 2018) can distinguish ADHD and ASD cases from controls in this sample. Findings from genetic studies suggest a partly shared genetic diathesis underlying neurodevelopmental disorders (including SCZ, ASD and ADHD) (Bulik-Sullivan et al. 2015). We therefore hypothesized that the ADHD, ASD and SCZ PRS would be associated with the ADHD/ASD (either ASD, ADHD or both) diagnostic status. In addition, we expected both the ADHD and ASD PRS to be associated with ADHD and ASD respectively. In addition, we expected the SCZ PRS to be associated with ASD status given the genetic overlap previously reported (Autism Spectrum Disorders Working Group of The Psychiatric Genomics Consortium 2017), although conflicting results with low (Cross-Disorder Group of the Psychiatric Genomics Consortium et al. 2013) or no (Vorstman et al. 2013) genetic association between ASD and SCZ have been reported as well. As a sensitivity analysis, we aim to perform a follow up correlation analysis and subsequently a linear regression analysis with the Child Behavioral Checklist (CBCL) subscales to validate the robustness of our findings and gain additional information on the link between associated genetic signals and specific behavioral or emotional problems, given a particular clinical diagnosis.
Methods
Sample
Psychiatric outpatient sample: “Inside-Out”
A new psychiatric outpatient sample called “Inside-Out” is analyzed. Data were collected from January 2001 until January 2012 at the department of Child and Adolescent Psychiatry at the Sophia Children’s Hospital at Erasmus Medical Center in Rotterdam, resulting in a psychiatric outpatient sample. Before the first visit, parents and children received the CBCL from the Achenbach System of Empirically Based Assessment (ASEBA) (Achenbach and Rescorla 2001). In addition, DNA was extracted from saliva and genotyping was performed on the Illumina PsychChip array (see data). The procedure was approved by the ethical committee of the Erasmus Medical Center. The total Inside-Out sample comprises 1941 children diagnosed with one or more DSM-IV disorders (ASD, ADHD, Tic disorder, Obsessive Compulsive Disorder (OCD), Depression, Anxiety, Anorexia Nervosa (AN), eating disorder NOS, RETT syndrome and subcategories of mentioned disorders) and children with a delayed diagnostic status or children who did not receive a DSM diagnosis (27.9%). The diagnostic procedure consisted of an interview with parents, a semi-structured interview with the child based on the Semi-structured Clinical Interview for Children and Adolescents (McConaughy and Achenbach 2001), the Diagnostic Interview Schedule for Children IV-P (Shaffer et al. 2000) and the Autism Diagnostic Observation Schedule-Generic (Lord et al. 1989) in case of a suspected autism spectrum disorder. Diagnostic classification was done by a clinician according to the Diagnostic and Statistical Manual of Mental Disorders, fourth edition (DSM-IV). The above-mentioned procedure was part of standard clinical practice For the current study, genetic and clinical information was used of the children who received an ADHD diagnosis, no ASD co-diagnosis allowed (N = 280, age range: 3.3–18.5 years, mean: 9.06, SD: 2.66) or an ASD diagnosis, no ADHD co-diagnosis allowed (RETT excluded) (N = 295, age range: 2.5–18.3 years, mean: 9.02, SD: 3.55). In addition we used a sample of combined ADHD and ASD diagnoses where comorbidity of ADHD and ASD was allowed, adding another 113 children to this combined sample (N = 688, age range 2.5–18.5, Mean: 8.96, SD: 3.07). The target sample was diagnosed with the DSM-IV and includes many cases with Asperger and pervasive developmental disorder-not otherwise specified (PDD-NOS) diagnoses (82% of total ASD sample). ADHD and ASD co-diagnosed children (N = 113) were not included in the ADHD and ASD sample. For sample specifics see Tables 1 and 2.
Population-based control sample
As a control sample, we used a Dutch population sample (NESCOG, N = 943, age range: 17.0–79.0) previously described by Polderman et al. (2013). NESCOG comprises a general population and a family-based sample of which closely related individuals were excluded. Data were collected on cognitive tasks, behavioral conditions (such as ADHD and ASD symptoms), life events, personality and environmental factors, as well as genetic information. Moreover, to correct for undiagnosed ADHD status, participants scoring 3 SD above the mean on the Conners’ Adult ADHD Rating Scale (CAARS) (Conners et al. z.d.), or the Attention Problems scale of the Young Adult Self Report (YASR) (Achenbach 1997) were excluded. Participants scoring three SD above mean on the Autism Quotient (AQ) (Baron-Cohen et al. 2001) were also excluded, resulting in a final control sample of 943 participants (age range 17–79, 38% male), see Tables 1 and 2.
Data
Genotyping of the cases and controls was performed on the same Illumina PsychChip array. The PsychChip SNP array contains HumanCore, Human Exome and custom content to accurately capture genetic variants previously linked with psychiatric disorders (https://www.illumina.com/products/by-type/microarray-kits/infinium-psycharray.html). Genetic variants in the clinical sample were filtered based on minor allele frequency (MAF < 1%), Hardy–Weinberg disequilibrium (P < 1 × 10−6) and SNP call rate (< 95%). Individuals were subsequently filtered based on relatedness (pairwise Identity-By-Descent (IBD) > 0.185), genotype and phenotypic sex mismatch, outlying heterozygosity and non-European ancestry (4 SD outside the range of the first two genetic principal components of the HapMap3 European founder population (CEU)) resulting in a clinical sample of 812 patients of which 688 are diagnosed with ADHD, ASD or both. The remaining part of the children in this sample (N = 124) are diagnosed with either Rett syndrome, Anorexia Nervosa or other eating disorders, Tourette Disorder, or other disorders. Another subset of the sample is currently being genotyped and includes children diagnosed with Anxiety Disorder, Affective Disorder or other disorders. In the control sample, SNP filtering was based on MAF (< 1%) Hardy–Weinberg disequilibrium (P < 0.00001) and SNP call rate (< 95%). Individual QC was based on missingness (> 5%), ancestry (within the range of 1000G CEU population on first PCs), relatedness (pairwise IBD > 0.185), gender mismatch, outlying heterozygosity and missing phenotypes.
Sex differences in the samples
The case and control samples differed in sex distribution (cases are 75% and the controls 25% males). Therefore, we compared allele frequencies between males and females in an independent sample, GoNL (see www.nlgenome.nl for more information), by means of correlation. The Pearson correlation coefficient between the male and female allele frequencies is 0.99, removing concerns of different allele frequencies in the two samples due to sex differences.
Polygenic risk scoring
The PRS is constructed as the sum of risk alleles weighted by their effect size. Per disorder several PRS were calculated with different P value inclusion thresholds (P-values: < 0.01, < 0.05, < 0.1, < 0.2, < 0.3, < 0.4, < 0.5, < 1). Starting from a low P-value threshold moving up to P-value 1, an optimal P-value threshold with the highest explained variance was identified, including the most truly associated positives. After this threshold more false positives will be included dampening the true signal. (Wray et al. 2014). Prior to our calculation of the PRS, the SNPs were pruned (LD R2 < 0.1, 250 kb pair window) to remove variants in LD. Polygenic scoring was performed with the software package PRSice (Euesden et al. 2015). The PRS for ASD, ADHD and SCZ were constructed using the most recent summary statistics from GWAS with the largest publicly available sample size, ADHD (Demontis and Walters 2017) (20,183 cases and 35,191 controls), ASD (Grove et al. 2017) (18,382 cases and 27,969 controls), and SCZ (Pardiñas et al. 2018) (40,675 cases and 64,643 controls). Of note, the Inside-Out and the control sample are independent samples, not included in these GWAS. After polygenic scoring the results were standardized to mean 0 and SD 1 for interpretational purposes. For the number of SNPs included in the scores see Supplementary Table S1.
Behavioral measurements
Child emotional and behavioral problems were assessed using the Dutch version of the Child Behavior Checklist/6–18 (CBCL) (Achenbach and Rescorla 2001) filled out by the parent before the first visit to the hospital. The CBCL contains 113 problem items that can be scored on eight syndrome scales (Anxious/Depressed Nitem = 13, Withdrawn/Depressed Nitem = 8, Somatic Complaints Nitem = 11, Social Problems Nitem = 11, Thought Problems Nitem = 15, Attention Problems Nitem = 10, Rule Breaking Behavior Nitem = 17 and Aggressive Behavior Nitem = 18). Parents score each problem on a three-point scale (0: not true, 1: somewhat or sometimes true, 2: very or often true). This follow up analysis included children with a CBCL 6–18 report, completed by the parent less than a year before receiving the diagnosis. If a CBCL from within a year before diagnosis was not present the person was excluded from this part of the analysis. In all analyses, sum scores on the CBCL syndrome scales were used.
Statistical analysis
Case control analysis on the association between PRS and disease status
We performed logistic regression analyses to investigate if the ADHD, ASD or SCZ PRS can distinguish between cases and controls in a sample (1) with a diagnosis of ADHD, ASD not permitted as co-diagnosis (ADHD, N = 280), (2) with a diagnosis of ASD, ADHD not permitted as co-diagnosis (ASD, N = 295), and (3) combining the first two samples, thus subjects with either ASD, ADHD or both (ADHD/ASD, N = 688). For each PRS, eight different SNP inclusion thresholds were tested. All P-values were corrected for multiple testing by means of Bonferroni correction (72 tests: three samples (ADHD, ASD, ADHD/ASD), three PRS (ADHD, ASD, SCZ), eight PRS thresholds (0.01, 0.05, 0.1, 0.2, 0.3, 0.4, 0.5, 1) per disorder). To account for population stratification we included eight principal components (PCs). The PCs were calculated based on the pruned data with Eigensoft (Price et al. 2006) (version 3.0) software. Additionally, sex was added as a covariate. Age was not added as a covariate as all cases are children and all controls are adults.
Sensitivity analysis: correlation and association between CBCL syndrome scales and PRS
We aim to provide additional evidence for the significant association of the PRS and the disorders as measured by the CBCL score severity. Given statistical power, we tested the association with symptom severity only in the combined ADHD/ASD sample by calculating the correlation between the significantly associated PRS (i.e., ADHD) and the syndrome scales of the CBCL. Age was added as a covariate in addition to the previously used eight PCs and sex. All analyses were performed in IBM SPSS statistics 21.
Results
Case control analysis on the association between PRS and disorder status
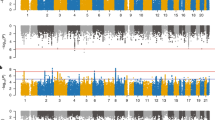
The ADHD PRS showed significant associations before multiple testing correction with disorder status in all three samples (Table 3). As shown in Fig. 1, all ADHD PRS P-value thresholds remained significant after Bonferroni multiple testing correction in both the combined ADHD/ASD and ADHD sample, but not the ASD sample. The most stringent P-value threshold of 0.01 generated a positive association in the ADHD/ASD sample OR 1.28 (P = 1.3 × 10−3), and ADHD sample OR 1.4 (P = 3.6 × 10−4). The most optimal P-value threshold as defined by explained variance, OR and P-value was 0.3 for the ADHD/ASD sample (R2 = 0.02, OR 1.36, P = 1.21 × 10−05), and 0.4 for the ADHD sample (R2 = 0.045, OR 1.62, P = 5.75 × 10−08).

Variance explained (Nagelkerke R2) by the ADHD PRS. All SNP inclusion P-value thresholds are shown from low to high (0.01, 0.05, 0.1, 0.2, 0.3, 0.4, 0.5, 1). P-values are shown on top of each bar and are Bonferroni corrected
The most lenient P-value threshold of P < 1 had a significant association in the ADHD/ASD sample, OR 1.35 (P = 1.9 × 10−5), and also in the ADHD sample OR 1.62 (P = 4.73 × 10−8). In the ASD sample none of the results remained significant after Bonferroni correction.
The ASD and SCZ PRS were not significantly associated with the ADHD, ASD, or combined ADHD/ASD status. The SCZ PRS including all SNPs (P-value threshold P < 1) showed a trend towards association in the ADHD/ASD sample (OR 1.13, P = 5.72 × 10−2) (Supplementary Tables S2 and S3 and Figs. S1 and S2).
Sensitivity analysis: association between CBCL syndrome scales and the ADHD PRS
Based on the correlational structure in the ADHD/ASD sample (Supplemental Material Table 4) between the CBCL syndrome scale scores and the ADHD PRS P-value thresholds, we concluded the correlation was too low (all correlations ≤ 0.1) to warrant the linear regression analysis. Mean scores and standard deviations for the CBCL syndrome scale scores for the ADHD/ASD sample are provided in Supplementary Table 5.
Discussion
This study investigated the associations of PRS for ADHD, ASD and SCZ, with ADHD and ASD status in a clinical child and adolescent population. As hypothesized, we found a significant association between the ADHD PRS and the combined ADHD/ASD status, and the separate ADHD status. The PRS SNP-inclusion thresholding resulted in a rise of explained variance with increasing P-value thresholds, showing that in addition to the GWAS significant hits, the non-significant SNPs in the ADHD GWAS also contribute to the associations with diagnostic status. Given the comorbidity between ADHD and ASD, and previously reported genetic correlations, we expected that the ADHD PRS would also be associated with ASD status, however, this association was not observed in our data. In contrast, the current results suggest a disorder specific effect of ADHD associated SNPs instead of a shared common genetic mechanism with ASD. The ADHD PRS is based on the most recent GWAS results, and explained variance up to 4.5% in our sample, which is in line with the results from the initial GWAS (Demontis and Walters 2017) who reported an explained variance of 5.5%, making it a promising PRS for further use in research on ADHD.
Contrary to our expectation, the ASD and SCZ PRS were not associated with any of the diagnostic groups. The null results for the ASD PRS are unexpected as the initial GWAS (Grove et al. 2017) reported an explained variance of 2.45% in an independent sample, and their summary statistics were used for the analysis. Given that the discovery sample size of ASD was only slightly smaller than the ADHD sample, and the SCZ sample was even larger, we do not expect that sample size alone explains these findings. Moreover, apart from sample size, power analyses usually take several parameters into account, including the heritability and population prevalence of traits, the amount of SNPs included in the GWAS, the effective number of chromosome segments, and the proportion of cases in discovery and target sample (Lee et al. 2017). In our study, the discovery and target samples were for most of the parameters similar across disorders, except for prevalence rates (ASD and SCZ have a population prevalence of 1%, and ADHD has a population prevalence of 5%).
Regarding the null result for the ASD PRS one explanation might be a difference in the diagnostic sample composition of the ASD GWAS discovery sample compared to the target ASD sample. The target sample was diagnosed with the DSM-IV and includes many cases with Asperger, and PDD-NOS diagnoses (82% of total ASD sample), which might differ from the discovery sample. Moreover, about one-third of the discovery sample were trio data (i.e. case pseudo control design), of which it has been suggested that the un-transmitted chromosomes contain increased polygenic burden, and as such the genetic signal based on these data might be decreased (Peyrot et al. 2016). Additionally, the genetic architecture of ADHD might differ from ASD, e.g., rare genetic variants might comprise a more important part of the genetic contribution to ASD (Geschwind and State 2015) compared to ADHD. With growing sample sizes, genetic discoveries will increase and become more reliable, potentially allowing the identification of rare variants.
The choice of including the SCZ PRS was based partly on the higher prevalence rate of SCZ in ASD individuals compared to the general population, a recent systematic review reports a significantly higher SCZ prevalence in ASD individuals compared to the general population (OR 3.55, 95% confidence interval (CI) 2.08–6.05, P < 0.001) (Zheng et al. 2018). If the actual SCZ prevalence rate in an ASD population resides at the lower end of the of the 95% CI the enrichment of common SCZ SNPs might not be detectable in our relatively small sample. Additionally, the genetic correlation of 0.211 between ASD and SCZ (Grove et al. 2017) and 0.122 between SCZ and ADHD (Demontis and Walters 2017) might be too small to detect the genetic overlap between the two disorders in our data. Finally, it is possible that ASD has a different genetic underpinning with more rare variants than SCZ although some overlap has been reported in rare genetic variation between ASD and SCZ (Sanders et al. 2015). Recent whole-genome sequence research on height fully recovered the heritability of this trait, meaning that next to the previously established common variants, all rare variants have been discovered (Wainschtein et al. 2019). Whole-genome sequence research into ASD, SCZ and ADHD might shed light on this issue revealing the genetic architecture of these traits.
The sensitivity analyses exploring the associations between scores on the syndrome scales of the CBCL and the ADHD PRS showed low correlations between these two measures, as such we decided not to pursue the follow-up analysis further. A reason for the low correlations can be the amount variance explained by the ADHD PRS. The explained variance of 4.5% might not be enough to give meaningful results in follow-up analysis using the CBCL in a smaller sample like “Inside out”. In addition, a diagnosis is not based solely on the CBCL results but includes careful evaluation by an experienced psychologist/psychiatrist based on a patient interview, a parent interview and if possible an evaluation by a third party like a school teacher of the child.
Strengths and limitations
A strength of our study is the adult control sample as, in contrast to a child sample, the chance that adult individuals will receive a future ADHD or ASD diagnosis is limited compared to young individuals i.e., these disorders are usually diagnosed during childhood (Nylander et al. 2013), while DNA sequences are fixed during life.
One concern might be the difference in sex distribution between the samples, with the clinical sample consisting of 75% males and the control sample having an opposite skew in sex distribution, as this could potentially affect the observed associations between the PRS and diagnoses. However, we compared the allele frequencies between males and females in an independent sample (GoNL (Genome of the Netherlands Consortium 2014)) and found no differences. Yet, due to the skewed sex distribution we could not examine sex-PRS interactions, or sex specific associations, which would both be interesting to investigate given the higher prevalence of males in both ADHD and ASD.
We also need to take into account that the ADHD/ASD group comprises the ADHD and ASD groups and that this is no official diagnostic disorder classification. The results should be replicated in a comparable independent sample first before firm conclusions can be drawn.
Overall, despite the fact that symptoms overlap between the neurodevelopmental disorders, our study does not directly imply that the umbrella of NDD is present at the common genetic level as captured in the PRS. As the ASD and SCZ PRS do not distinguish cases from controls in any of our diagnostic samples it is possible that ADHD, ASD and SCZ have a different common genetic signature. Moreover, the results should be replicated in one or more independent samples.
A final remark can be made on the cross sectional nature of the sample. Unlike longitudinal studies, measures are available for one point in time for most of the subjects. This presents the possibility that children might receive additional diagnoses later on in life resulting in a change in diagnostic status from ADHD or ASD to the ADHD/ASD codiagnosed group, or to other comorbidities.
Conclusions
In conclusion, the PRS of ADHD is significantly associated with the combined ADHD/ASD and ADHD status. Yet, this association is primarily driven by ADHD status, suggesting disorder specific genetic effects of the ADHD PRS. Nevertheless, it is of interest to explore the genetic predictive value of other psychiatric disorders besides neurodevelopmental disorders. Improving genetic prediction in neurodevelopmental disorders by using a multi-trait predictor instead of single-trait predictors is also an interesting option (Maier et al. 2018). Lastly, it is of interest to delve deeper into the association between the ADHD PRS and the specific emotional and behavioral problems in larger samples as those data may provide additional information on specific problems or the severity of problems within a diagnostic status.
References
Achenbach TM (1997) Manual for the young adult self-report and young adult behavior checklist. Department of Psychiatry, University of Vermont, Burlington
Achenbach TM, Rescorla LA (2001) Manual for the ASEBA school-age forms & profiles. University of Vermont, Research Center for Children, Youth, & Families, Burlington
American Psychiatric Association (2013) Diagnostic and statistical manual of mental disorders, vol 5. American Psychiatric Association, Washington, DC
Autism Spectrum Disorders Working Group of The Psychiatric Genomics Consortium (2017) Meta-analysis of GWAS of over 16,000 individuals with autism spectrum disorder highlights a novel locus at 10q24.32 and a significant overlap with schizophrenia. Mol Autism 8:21. https://doi.org/10.1186/s13229-017-0137-9
Baron-Cohen S, Wheelwright S, Skinner R, Martin J, Clubley E (2001) The autism-spectrum quotient (AQ): evidence from Asperger syndrome/high-functioning autism, males and females, scientists and mathematicians. J Autism Dev Disord 31(1):5–17
Brikell I, Kuja-Halkola R, Larsson H (2015) Heritability of attention-deficit hyperactivity disorder in adults. Am J Med Genet B 168(6):406–413. https://doi.org/10.1002/ajmg.b.32335
Bulik-Sullivan B, Finucane HK, Anttila V, Gusev A, Day FR, Loh P-R et al (2015) An atlas of genetic correlations across human diseases and traits. Nat Genet. https://doi.org/10.1038/ng.3406
Cheung C, Yu K, Fung G, Leung M, Wong C, Li Q et al (2010) Autistic disorders and schizophrenia: related or remote? An anatomical likelihood estimation. PLoS ONE 5(8):e12233. https://doi.org/10.1371/journal.pone.0012233
Conners CK, Erhardt D, Sparrow E (z.d.). Conner’s adults ADHD rating scales. North Tonawanda, New York
Craig F, Lamanna AL, Margari F, Matera E, Simone M, Margari L (2015) Overlap between autism spectrum disorders and attention deficit hyperactivity disorder: searching for distinctive/common clinical features. Autism Res 8(3):328–337. https://doi.org/10.1002/aur.1449
Cross-Disorder Group of the Psychiatric Genomics Consortium, Lee SH, Ripke S, Neale BM, Faraone SV, Purcell SM et al (2013) Genetic relationship between five psychiatric disorders estimated from genome-wide SNPs. Nat Genet 45(9):984–994. https://doi.org/10.1038/ng.2711
Demontis D, Walters RK (2017) Discovery of the first genome-wide significant risk loci for ADHD. Nat Genet. https://doi.org/10.1101/145581
Demontis D, Walters RK, Martin J, Mattheisen M, Als TD, Agerbo E, Baldursson G, Belliveau R, Bybjerg-Grauholm J, Bækvad-Hansen M et al (2019) Discovery of the first genome-wide significant risk loci for attention deficit/hyperactivity disorder. Nat Genet 51:63–75
Euesden J, Lewis CM, O’Reilly PF (2015) PRSice: polygenic risk score software. Bioinformatics 31(9):1466–1468. https://doi.org/10.1093/bioinformatics/btu848
Genome of the Netherlands Consortium (2014) Whole-genome sequence variation, population structure and demographic history of the Dutch population. Nat Genet 46(8):818–825. https://doi.org/10.1038/ng.3021
Geschwind DH, State MW (2015) Gene hunting in autism spectrum disorder: on the path to precision medicine. Lancet Neurol 14(11):1109–1120. https://doi.org/10.1016/S1474-4422(15)00044-7
Gratten J, Wray NR, Keller MC, Visscher PM (2014) Large-scale genomics unveils the genetic architecture of psychiatric disorders. Nat Neurosci 17(6):782–790. https://doi.org/10.1038/nn.3708
Groen-Blokhuis MM, Middeldorp CM, Kan K-J, Abdellaoui A, van Beijsterveldt CEM, Ehli EA et al (2014) Attention-deficit/hyperactivity disorder polygenic risk scores predict attention problems in a population-based sample of children. J Am Acad Child Adolesc Psychiatry 53(10):1123–1129.e6. https://doi.org/10.1016/j.jaac.2014.06.014
Grove J et al (2017) Common risk variants identified in autism spectrum disorder. bioRxiv. https://doi.org/10.1101/224774
Grove J, Ripke S, Als TD, Mattheisen M, Walters RK, Won H, Pallesen J et al (2019) Identification of common genetic risk variants for autism spectrum disorder. Nat Genet 51(3):431–444. https://doi.org/10.1038/s41588-019-0344-8
Jansen PR, Polderman TJC, Bolhuis K, van der Ende J, Jaddoe VWV, Verhulst FC et al (2017) Polygenic scores for schizophrenia and educational attainment are associated with behavioural problems in early childhood in the general population. J Child Psychol Psychiatry. https://doi.org/10.1111/jcpp.12759
Lee SH, Weerasinghe WMSP, Wray NR, Goddard ME, van der Werf JHJ (2017) Using information of relatives in genomic prediction to apply effective stratified medicine. Sci Rep 7:42091. https://doi.org/10.1038/srep42091
Lord C, Rutter M, Heemsbergen J, Jordan H, Mawhood L, Schopler E (1989) Autism diagnostic observation schedule: a standardized observation of communicative and social behavior. J Autism Dev Disord 19(2):185–212
Maier RM, Zhu Z, Lee SH, Trzaskowski M, Ruderfer DM, Stahl EA et al (2018) Improving genetic prediction by leveraging genetic correlations among human diseases and traits. Nat Commun 9(1):989. https://doi.org/10.1038/s41467-017-02769-6
Martin J, Hamshere ML, Stergiakouli E, O’Donovan MC, Thapar A (2014) Genetic risk for attention-deficit/hyperactivity disorder contributes to neurodevelopmental traits in the general population. Biol Psychiatry 76(8):664–671. https://doi.org/10.1016/j.biopsych.2014.02.013
McConaughy SH, Achenbach TM (2001) Manual for the semi-structured clinical interview for children and adolescents. Department of Psychiatry, University of Vermont, Burlington
Mitchell KJ (2011) The genetics of neurodevelopmental disease. Curr Opin Neurobiol 21:197–203
Mullin AP, Gokhale A, Moreno-De-Luca A, Sanyal S, Waddington JL, Faundez V (2013) Neurodevelopmental disorders: mechanisms and boundary definitions from genomes, interactomes and proteomes. Transl Psychiatry 3(12):e329. https://doi.org/10.1038/tp.2013.108
Nylander L, Holmqvist M, Gustafson L, Gillberg C (2013) Attention-deficit/hyperactivity disorder (ADHD) and autism spectrum disorder (ASD) in adult psychiatry. A 20-year register study. Nord J Psychiatry 67(5):344–350. https://doi.org/10.3109/08039488.2012.748824
Pardiñas AF, Holmans P, Pocklington AJ, Escott-Price V, Ripke S, Carrera N et al (2018) Common schizophrenia alleles are enriched in mutation-intolerant genes and in regions under strong background selection. Nat Genet 50(3):381–389. https://doi.org/10.1038/s41588-018-0059-2
Peyrot WJ, Boomsma DI, Penninx BWJH, Wray NR (2016) Disease and polygenic architecture: avoid trio design and appropriately account for unscreened control subjects for common disease. Am J Hum Genet 98(2):382–391. https://doi.org/10.1016/j.ajhg.2015.12.017
Polderman TJC, Hoekstra RA, Vinkhuyzen AAE, Sullivan PF, van der Sluis S, Posthuma D (2013) Attentional switching forms a genetic link between attention problems and autistic traits in adults. Psychol Med 43(9):1985–1996. https://doi.org/10.1017/S0033291712002863
Price AL, Patterson NJ, Plenge RM, Weinblatt ME, Shadick NA, Reich D (2006) Principal components analysis corrects for stratification in genome-wide association studies. Nat Genet 38(8):904–909. https://doi.org/10.1038/ng1847
Rapoport JL, Giedd JN, Gogtay N (2012) Neurodevelopmental model of schizophrenia: update 2012. Mol Psychiatry 17(12):1228–1238. https://doi.org/10.1038/mp.2012.23
Sanders SJ, He X, Willsey AJ, Ercan-Sencicek AG, Samocha KE, Cicek AE et al (2015) Insights into autism spectrum disorder genomic architecture and biology from 71 risk loci. Neuron 87(6):1215–1233. https://doi.org/10.1016/j.neuron.2015.09.016
Shaffer D, Fisher P, Lucas CP, Dulcan MK, Schwab-Stone ME (2000) NIMH Diagnostic Interview Schedule for Children Version IV (NIMH DISC-IV): description, differences from previous versions, and reliability of some common diagnoses. J Am Acad Child Adolesc Psychiatry 39(1):28–38. https://doi.org/10.1097/00004583-200001000-00014
Sullivan PF, Kendler KS, Neale MC (2003) Schizophrenia as a complex trait: evidence from a meta-analysis of twin studies. Arch Gen Psychiatry 60(12):1187–1192. https://doi.org/10.1001/archpsyc.60.12.1187
Sullivan PF, Daly MJ, O’Donovan M (2012) Genetic architectures of psychiatric disorders: the emerging picture and its implications. Nat Rev Genet 13(8):537–551. https://doi.org/10.1038/nrg3240
Tick B, Bolton P, Happé F, Rutter M, Rijsdijk F (2016) Heritability of autism spectrum disorders: a meta-analysis of twin studies. J Child Psychol Psychiatry 57(5):585–595. https://doi.org/10.1111/jcpp.12499
Torkamani A, Wineinger NE, Topol EJ (2018) The personal and clinical utility of polygenic risk scores. Nat Rev Genet. https://doi.org/10.1038/s41576-018-0018-x
Visscher PM, Wray NR, Zhang Q, Sklar P, McCarthy MI, Brown MA, Yang J (2017) 10 years of GWAS discovery: biology, function, and translation. Am J Hum Genet 101(1):5–22. https://doi.org/10.1016/j.ajhg.2017.06.005
Vorstman JAS, Anney RJL, Derks EM, Gallagher L, Gill M, de Jonge MV et al (2013) No evidence that common genetic risk variation is shared between schizophrenia and autism. Am J Med Genet B 162B(1):55–60. https://doi.org/10.1002/ajmg.b.32121
Wainschtein P, Jain DP, Yengo L, Zheng Z et al (2019) Recovery of trait heritability from whole genome sequence data. bioRxiv. https://doi.org/10.1101/588020
Weiner DJ, Wigdor EM, Ripke S, Walters RK, Kosmicki JA, Grove J et al (2017) Polygenic transmission disequilibrium confirms that common and rare variation act additively to create risk for autism spectrum disorders. Nat Genet 49(7):978–985. https://doi.org/10.1038/ng.3863
Wray NR, Lee SH, Mehta D, Vinkhuyzen AAE, Dudbridge F, Middeldorp CM (2014) Research review: polygenic methods and their application to psychiatric traits. J Child Psychol Psychiatry 55(10):1068–1087. https://doi.org/10.1111/jcpp.12295
Zheng Z, Zheng P, Zou X (2018) Association between schizophrenia and autism spectrum disorder: a systematic review and meta-analysis. Autism Res 11(8):1110–1119. https://doi.org/10.1002/aur.1977
Acknowledgements
We would like to thank the PGC for allowing us to use their summary statistics. Statistical analyses were carried out on the Genetic Cluster Computer (http://www.geneticcluster.org), which is financially supported by the Netherlands Scientific Organization (NWO 480-05-003). We gratefully acknowledge financial support by the Sophia Stichting voor Wetenschappelijk Onderzoek under grant number 593 and S14-27, and the Netherlands Scientific Organization (NWO VICI 453-14-005).
Funding
The funding organizations had no role in any of the following: the design and conduct of the study; collection, management, analysis, and interpretation of the data; and preparation, review, or approval of the manuscript.
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
Conflict of interest
Frank Verhulst is the distributor of the Dutch translations of ASEBA from which he receives renumeration. Arija G. Jansen, Gwen C. Dieleman, Philip R. Jansen, Danielle Posthuma, and Tinca J. C. Polderman report no conflict of interest.
Human and animal rights and informed consent
The Erasmus MC medical committee approved all procedures in this study which is performed in accordance with the 1964 Helsinki declaration and it's later amendments or comparable ethical standards. As the subjects were under 18 years of age, legal guardians provided informed consent for the participants prior to inclusion and testing in this study.
Additional information
Handling Editor: Chandra A. Reynolds, PhD.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Jansen, A.G., Dieleman, G.C., Jansen, P.R. et al. Psychiatric Polygenic Risk Scores as Predictor for Attention Deficit/Hyperactivity Disorder and Autism Spectrum Disorder in a Clinical Child and Adolescent Sample. Behav Genet 50, 203–212 (2020). https://doi.org/10.1007/s10519-019-09965-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10519-019-09965-8