
Abstract
Superiority of the left upper limb in proprioception tasks performed by right-handed individuals has been attributed to better utilization of proprioceptive information by a non-preferred arm/hemisphere system. However, it is undetermined whether this holds for multiple upper and lower limb joints. Accordingly, the present study tested active movement proprioception at four pairs of upper and lower limb joints, after selecting twelve participants with both strong right arm and right leg preference. A battery of versions of the active movement extent discrimination apparatus were employed to generate the stimuli for movements of different extents at the ankle, knee, shoulder and fingers on the right and left sides of the body, and discrimination scores were derived from participants’ responses. Proprioceptive performance on the non-preferred left side was significantly better than the preferred right side at all four joints tested (overall F 1, 11 = 36.36, p < 0.001, partial η 2 = 0.77). In the 8 × 8 matrix formed by all joints, only correlations between the proprioceptive accuracy scores for the right and left sides at the same joint were significant (ankles 0.93, knees 0.89, shoulders 0.87, fingers 0.91, p ≤ 0.001; all others r ≤ 0.40, p ≥ 0.20). The results point to both a side-general effect and a site-specific effect in the integration of proprioceptive information during active movement tasks, whereby the non-preferred limb/hemisphere system is specialized in the utilization of the best proprioceptive sources available at each specific joint, but the combination of sources employed differs between body sites.
Similar content being viewed by others


Introduction
Following early limb positioning studies (Lloyd and Caldwell 1965; Paillard and Brouchon 1968), recent studies by Goble and colleagues have explored asymmetries in upper limb proprioception, using both passive and active position matching paradigms, which also require interhemispheric transfer or memory retrieval of proprioceptive information at the elbow (Goble et al. 2006; Goble and Brown 2007, 2008b, 2009, 2010). These studies have demonstrated a non-preferred arm/hemisphere system superiority in the utilization of proprioceptive feedback in right-handed individuals (Goble et al. 2006). However, all these results were obtained from a single upper limb joint, the elbow, so it is not known if this effect is evident at other upper and lower limb joints, such as the shoulders, fingers, knees and ankles. A question therefore remains as to whether non-preferred limb/hemisphere system specialization in proprioceptive tasks is a side-general attribute or a site-specific attribute.
Non-preferred arm/hemisphere specialization in the utilization of proprioceptive feedback has been attributed to functional differences between the roles of the preferred and non-preferred arms in bimanual tasks (Goble et al. 2009). That is, the role of the non-dominant arm is to statically stabilize an object for the dominant arm to dynamically manipulate, and this division of functions has conferred a non-preferred arm/hemisphere superiority in the utilization of proprioceptive feedback. Such a notion is consistent with Sainburg’s proposition that the non-preferred arm/hemisphere system is specialized for static limb position control, whereas the preferred arm/hemisphere system is specialized for dynamic limb trajectory control (Sainburg 2002, 2005). Thus, during bilateral tasks such as removing a lid, or kicking a ball, joints on the non-dominant side usually work in concert to stabilize an object, or stabilize the body, for joints on the dominant side to manipulate. According to this view, all joints on the non-dominant side would be expected to perform better on position-related proprioception tasks.
However, proprioceptive asymmetry may be evident only at distal joints, not at proximal joints. It has been argued that whereas proximal musculature has been thought to be innervated by both hemispheres, more distal musculature has been considered to be innervated largely by the contralateral hemisphere (Kuypers 1982; Muller et al. 1991). Supporting this notion, an early classical study by Roy and MacKenzie (1978) explored the accuracy of individuals when matching thumb and multi-joint arm positions without visual cues. Their results showed non-preferred left limb superiority for matching thumb position (distal), while no side differences were evident in terms of arm position matching (proximal). Similarly, in lower limb studies where knee joint position sense was measured under non-weight-bearing, partial weight-bearing and full weight-bearing conditions, the results showed no side differences in any condition (Bullock-Saxton et al. 2001). In contrast, another study assessed more distal movement discrimination ability at both ankles across three inversion extents and found the non-preferred left ankles to be significantly better at all three inversion depths (Symes et al. 2010). Work is thus needed to systematically test both upper and lower limbs at multiple proximal and distal joints, in order to determine whether proprioceptive asymmetry is a side-general attribute that is evident at different joints around the body.
When exploring proprioceptive ability at different joints, another important question is whether ability to utilize proprioceptive information at different joints is a body-general attribute or a site-specific attribute, that is, whether subgroups that perform better/worse on a proprioceptive task at one joint are also those who perform better/worse at other joints.
It has been suggested that joint injury may be more likely when there is a “pre-existing global deficit” in proprioception (Goldie et al. 1994). That is, for those with such a deficit, utilization of proprioceptive information may be generally poor at different sites around the body. Therefore, if the relatively poor performers on proprioceptive tasks are those with low ability to integrate information from different sources, they will maintain their low ranking across different testing sites, and as a consequence, there will be significant correlations between the scores obtained from different body sites.
Contrary evidence, however, suggests that ability to use proprioception at different joints may not be a body-general attribute, but rather that proprioceptive ability at different joints is site-specific. If proprioception at different joints is experience-dependent, then it would be expected that athletes whose sports skills were largely based on use of a specific body part would show significantly greater site-specific proprioceptive acuity. Consistent with this account, Muaidi et al. (2009) found significantly better knee rotation movement discrimination ability in Olympic-level soccer players than non-athletes. Thus, proprioceptive acuity is not significantly correlated across different joints with differing patterns of use. In their study of movement extent discrimination made without vision at the ankle and knee, Waddington and Adams (1999a) who also used athletes found significant correlations only between the movement discrimination scores at the two ankles and the two knees, but not between the ankles and knees on the same side. This study was, however, limited to two joints within the lower limb, and it has not been determined whether other joints in the upper limbs follow the same pattern.
It has been argued that in testing proprioceptive acuity, it is important that the tests maximize the similarity between the laboratory test and real life function, that is, maximize ecological validity (Gibson 1979). However, there is a trade-off between maximizing ecological validity and maximizing experimental control, in that things that increase one typically reduce the other. Thus, use of straps to isolate the testing body parts, and using goggles and earmuffs to occlude vision and audition, ensures that these modalities are not available, but decreases the similarity of the test to normal function, and consequently limits generalizability (Ashton-Miller 2000). Therefore, the current study set out to maximize ecological validity, but also to control important experimental variables, so that individuals could integrate all normally available proprioceptive information from different receptors, such as cutaneous receptors, joint receptors and muscle spindles at different joints, during task performance. We employed a battery of versions of the active movement extent discrimination apparatus (AMEDA) (Waddington and Adams 1999b; Waddington et al. 2000; Naughton et al. 2005; Symes et al. 2010; Han et al. 2011) that all involve active movements, made under normal weight-bearing conditions, without physical constraints, and all permit general vision while obscuring vision of the target position. In addition, the nature of AMEDA tasks ensures that information about both limb movement extent and end position is available on every trial, and this combination allows for better performance than that which is seen with extent information alone (Magill and Parks 1983).
By examining active movement discrimination at the left and right ankles, left and right knees, left and right shoulders and left and right finger and thumb digits of individuals with a strong right side preference, we sought to determine whether there is a side-general (both proximal and distal joints) or site-specific characteristic of proprioceptive asymmetry between the right and left sides of the body, and whether there is a body-general or site-specific attribute, in terms of the relationship between different body sites. Specifically, if proprioceptive asymmetry is a side-general attribute, individuals with a strong right limb preference would be expected to show non-preferred left side superiority at all tested joints in both upper and lower limbs, while if proprioceptive asymmetry is a site-specific attribute, non-preferred left side superiority would be evident only at some joints. Further, if proprioceptive ability is a body-general attribute, then there should be a significant positive and high correlation between the measures of movement discrimination of all joints tested, while if proprioceptive ability is site-specific, then significant positive and high correlation should be observed only between the right and left sides at the same joints, with low and insignificant correlations between different joints.
Methods
Participants
Twelve healthy young volunteers (6 males and 6 females, mean age 21.4 years, SD ± 1.4; mean height 171.4 cm, SD ± 1.4; mean weight 60.3 kg, SD ± 7.6) were recruited from an advertisement placed on a university campus notice board. All potential participants were screened prior to inclusion with a health questionnaire to rule out any current or previous conditions that could affect proprioception, in particular: (1) presence of joint injuries within the past 6 months, (2) a diagnosis of chronic diseases (e.g. multiple sclerosis, stroke, Parkinson’s disease, rheumatoid arthritis, type 2 diabetes) (Wycherley et al. 2005) and 3) any history of visual or vestibular disturbance affecting balance (Waddington and Adams 2003; Kanegaonkar et al. 2012). All volunteering participants had no chronic disease, and no specific athletic, musical or other intensive motor skills training experience.
Only participants who demonstrated both strong right upper and strong lower limb preference were included. Handedness was measured using a ten-item version of the Edinburgh Handedness Inventory (Oldfield 1971). The mean ± SD laterality quotient score for participants was +96.25 ± 4.33; range +90 to +100, where scores of +100 represent an extreme right hander and scores of −100 represent an extreme left hander. Footedness was measured using a nine-item corrected version of the Foot Preference Inventory (Kalaycioglu et al. 2008). The mean ± SD laterality quotient score for participants was +0.42 ± 0.51; range 0 to +1, where a score of 0 represents an extreme right footer and a score of +9 represents an extreme left footer. Thus, all participants in the study were strongly right side dominant for both handedness and footedness.
The project was approved by the University of Canberra Committee for Ethics in Human Research (CEHR 11–47), and before the experimental session commenced, each participant signed an informed consent form, indicating their willingness to participate.
Apparatus
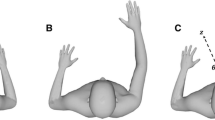
For each test, a purpose-built AMEDA, used in previous studies, was employed at four body sites: the ankle (Waddington and Adams 1999b; Waddington et al. 1999; Symes et al. 2010), knee (Waddington and Adams 1999a; Waddington et al. 2000), shoulder (Naughton et al. 2005; Vulcetic et al. 2008) and fingers (Han et al. 2011) (Fig. 1a). For the purpose of maximizing ecological validity, participants were not physically constrained or blindfolded in the AMEDA apparatus, and the psychophysical method employed has been shown to fulfil the validity criteria that have been proposed for assessing active movement function with sufficient trials to determine the ability to discriminate movement at different body sites (Ashton-Miller 2000).

a illustrates the four movement extent discrimination tasks for the ankle, knee, shoulder and fingers; b shows mean movement discrimination scores (AUC ± SD) for both sides of the body at each site (*p < 0.05)
Procedure
Participants were asked to wear a loose shirt, short pants and be barefoot to eliminate any possible confounders from clothing or footwear. In all tests, vision of the relevant body segment under investigation when it reached the physical stop under investigation was obscured, to ensure the judgement was based on proprioceptive information.
The procedures for AMEDA testing of active movements of ankle inversion, knee flexion, shoulder flexion and finger pinch discrimination have been described previously (Waddington et al. 2000; Naughton et al. 2005; Symes et al. 2010; Han et al. 2011). In brief, the AMEDA tests employed in the present study required participants to make active movements of ankle inversion, knee flexion, shoulder flexion or finger pinch to a physical stop, which can be varied, and the participant provided an absolute judgement of movement extent on return to the starting neutral position. That is, participants were asked to decide which one of the five possible movement distances they had just experienced, where there were five possible responses for each different movement distance. For each joint, a set of movement distance stimuli were selected to create a moderately difficult discrimination task.
Each participant was given a familiarization session on each test before data collection wherein they were informed that they would experience the five movement displacement distances, in order, from the smallest (moving to position 1) to the largest (moving to position 5), three times: 15 movements in total. Participants thereafter undertook 50 trials, without feedback, at each site. During each test set, trials were presented in a random sequence, 10 at each of the 5 different movement displacements. The order of testing of the two sides—right and left, and the four sites—ankle, knee, shoulder and fingers was also randomly assigned.
During the ankle AMEDA test, participants were instructed to stand astride, with the resting foot on the stable platform and the test foot centred over the axis of the swinging plate. While distributing weight evenly on both feet, with eyes looking forward at a point on the opposite wall, participants made an active ankle inversion movement at a steady pace from the horizontal fixed position, until the outer side rim of the moveable plate contacted the metal stepper motor rod. The plate was then returned to the start position at the same pace, where participants gave a judgment as to the extent of ankle inversion. The 5 predetermined ankle inversion displacements from smallest to largest were 10, 11, 12, 13 and 14 degrees.
For the knee AMEDA test, participants were instructed to stand in a lunge position facing towards to the height-adjustable knee AMEDA. The big toe of the testing leg was aligned with a tape marker on the ground, to ensure the same original testing position for both sides and that the greatest flexion movement of the knee would not pass the big toe. The knee AMEDA was then adjusted to the height of participant’s patella. While standing upright, with eyes looking forward at a point on the opposite wall, participants made an active knee flexion movement at a steady pace from a neutral standing position, until their patella touched the stepper motor plate. After returning to the upright position at the same pace, participants gave a judgment as to the extent of knee flexion. The 5 predetermined knee flexion displacements from smallest to largest were 37, 38, 39, 40 and 41 degrees.
In the shoulder AMEDA test, participants were instructed to stand astride, opposite the height-adjustable shoulder AMEDA. With one arm raised overhead and the back of the hand touching the moveable plate when in the zero position, a string with a plumb bob attached was used to obtain vertical alignment of the arm with the trunk. The heel position was then marked with tape on the ground for both sides, to preserve the original testing position. While standing upright, with the eyes looking forward to a wall 5 m away, the participants then made an active shoulder flexion movement at a steady pace from a neutral standing position with arms at the sides of the body, until the back of the hand touched the stepper motor plate. After returning the arm to the start position at the same pace, participants gave a judgment as to the extent of shoulder flexion. The 5 predetermined shoulder flexion displacements from smallest to largest were 170.6, 171.2, 171.7, 172.3 and 172.9 degrees.
During the finger pinch AMEDA test, participants sitting comfortably, with the thumb and index finger in place, were asked to make, at a steady pace, an active pinch movement that moved the tubes from a fixed start position dictated by the apparatus design closer to the midline until contact with the metal stop was made by the medial rims of the tubes. Participants returned the tubes to the start position and were asked to make a judgement as to the extent of pinch movement as soon as they returned the tubes to the start position. The 5 predetermined finger pinch displacements were obtained by measuring the distance from the end of the tube in the fixed start position to the testing position when the tube contacted the rim of the metal stop. These movement distances were from smallest to largest: 5.5, 8.0, 10.5, 13.0 and 15.5 mm.
Reliability
A reliability study was conducted with a 7-day interval between the sessions, with testing at the same time each day to minimize diurnal variations in joint laxity (Calguneri et al. 1982) using the procedures outlined above, but with a new randomized sequence. A single examiner carried out all experiments on all subjects, to eliminate any variability from utilizing separate observers (Jobbins et al. 1979).
Data analysis
For each test, the raw data were entered into a 5 × 5 matrix representing the frequency with which each response was made for each stimulus. Nonparametric signal detection analysis was applied to produce pair-wise receiver operating characteristic (ROC) curves, that is, comparing responses between distances 1–2, 2–3, 3–4 and 4–5 (McNicol 2004). Thereafter, the mean pair-wise area under the curve (AUC) was calculated using SPSS software V.18 to give each participant a single movement discrimination score. The AUC as a discrimination statistic provides an unbiased measure of ability to discriminate that is based on a signal detection theory approach to the sensory decision process (Swets et al. 1961, 2000; Swets 1996; McNicol 2004). AUC values range from 0.5, which is equivalent to chance responding, to 1.0, representing perfect ability to discriminate between the 5 different movement extents.
Reliability between repeated trials separated by 7 days was analysed using the interclass correlation coefficient (ICC). Where ICCs are high (Fleiss 1986), the data from two sessions can be averaged for further analysis. Repeated measures analysis of variance (ANOVA) was used to compare the data between joints of preferred and non-preferred sides to determine whether any difference existed between the same joints on either side of the body. Next, to determine whether the proprioception scores reflected the concept of proprioception as a body-general attribute or a site-specific attribute, Pearson correlations between the mean movement discrimination scores of the 4 sites—ankle (preferred/right and non-preferred/left), knee (preferred/right and non-preferred/left), shoulder (preferred/right and non-preferred/left) and fingers (preferred/right and non-preferred/left)—were determined. Finally, to obtain an overall test of preferred versus non-preferred side performance, a 2 × 4 × 2 factorial repeated measures ANOVA was conducted, with factors Preference (preferred, non-preferred), Joint (ankle, knee, shoulder, fingers) and Time (week one, week two). Cohen’s d (0.2 = small, 0.5 = medium, 0.8 = large) (Cohen 1992) was used to assess the size of effect associated with the experimental conditions.
Results
No significant test–retest difference was found between movement discrimination scores obtained at different sites of the body (ankle, p = 0.56; knee, p = 0.18; shoulder, p = 0.79; finger digits, p = 0.60) when tested 7 days apart, reflecting stability of the test scores. The ICCs for scores from testing 7 days apart at each site (ankle, knee, shoulder and fingers) were 0.96, 0.82, 0.86 and 0.88, respectively, all values which are in the high reliability range (Fleiss 1986). Given these results, data from the duplicate trials were averaged for each joint.
Mean movement discrimination scores with standard deviations were calculated and are reported in Table 1. ANOVA with repeated measures demonstrated a significant difference for movement discrimination between the preferred/right and non-preferred/left side of the body, with discrimination scores for the non-preferred left side significantly higher overall than for the preferred right side (F 1, 11 = 36.36, p < 0.001, partial η 2 = 0.77). Simple effects tests conducted on the movement discrimination scores at the ankles, knees, shoulders and fingers showed that the non-preferred/left side performed consistently better than the preferred/right side at all 4 paired joints (see Table 1 and Fig. 1b). There was no overall significant main effect of time of testing (F 1, 11 = 1.47, p = 0.25, partial η 2 = 0.12), nor any interaction effect involving time of testing (all p > 0.37).
The movement discrimination scores on the right and left sides for each participant at each joint are plotted in Fig. 2. The different ranges on the Y-axis reflect the fact that although all subjects could discriminate between the movements at the different joints to some extent, and not perfectly, the selection of movement extents to create a difficult, but not impossible, task at each joint produced a significant main effect of Joint (F 3, 33 = 78.75, p < 0.001). Cohen’s d for the overall side-of-the-body effect was calculated and found to be 0.72. With respect to the main aim of the study, comparing performance of the preferred and non-preferred sides on tasks that were equally difficult, the predominance of upward sloping lines reflects general left side superiority. The majority (79 %) of the side-to-side comparisons show left superiority. The effect was significant overall and at each joint. The relative performance of individuals on the four tasks can be examined by tracing the same symbol from left to right, over the four panels. For example, participant A can be seen to perform well, poorly and mid-range over the proprioceptive tasks at different body sites.

Movement discrimination scores of each participant for the ankles, knees, shoulders and fingers. Letters A–L represent the same participant’s performance on the different movement discrimination tasks. The different ranges on the Y-axis reflect differences in the difficulty of the four tasks
The matrix of Pearson correlation values is reported in Table 2. The different body sites showed significant positive and high correlations for movement discrimination only between the contralateral joints at a specific site. For correlations over different joints and sides, all r values were ≤0.40, p ≥ 0.20.
Discussion
Two important findings emerge from the current study. Firstly, for individuals with a strong preference for using their right limbs (both upper and lower), scores for discrimination of movement from the joints on the non-preferred/left side are consistently better using the left fingers, left shoulder, left knee and left ankle. Thus, the data demonstrate a side-general asymmetry, in terms of left side proprioceptive task superiority, at multiple upper and lower limb joints. Further, this left side superiority was evident for both proximal and distal joints in the upper and lower limbs. Secondly, Pearson correlations among movement discrimination scores of the tested joints showed significant positive and high correlations only between the right and left sides of the same joint, with low, non-significant correlations between different body sites, suggesting a site-specific attribute in the way that the brain integrates proprioceptive information from sources at different pairs of joints.
A strength of the current study is that it included multiple upper and lower limb joints, in comparison with research with one pair of joints, usually in the upper body (e.g. Nishizawa 1991; Goble et al. 2006; Adamo and Martin 2009). The finding that movement discrimination ability of right-handed/right-footed individuals is significantly better at multiple joints—their non-dominant left fingers, shoulder, knee and ankle—both extends and is consistent with previous findings obtained from a single joint (Goble et al. 2006; Goble and Brown 2009, 2010).
The differences between the AUC mean values for the joints tested on the left and right sides in the current study on average were 0.015. The AUC measure of discrimination sensitivity has been considered to be a reliable measure of ability to discriminate that is based on a signal detection theory approach to the sensory decision process (Swets et al. 1961, 2000; Swets 1996; McNicol 2004). The AUC is a proportion of the total area under the ROC curve (McNicol 2004), where the total area is set at 1.0, and chance performance on the discrimination test is 0.5, generating a restriction in range of AUC scores to between 0.5 and 1.0. A consequence of this is that the absolute differences between conditions that show significant differences in performance are small. However, the standardized mean difference, SMD, or Cohen’s d for the overall side-of-the-body main effect was 0.72, corresponding to a medium-large effect (1992). Thus, although the absolute values of the differences between the left and right sides were small, the associated effect size is a medium-large one.
Although small, differences in movement discrimination between left and right sides do have real-world significance. Waddington and Adams found that the difference between the mean AUC for ankle testing under barefoot or sport boot conditions was 0.017, and the effect of adding textured insoles was 0.021 (Waddington and Adams 2003). Error in discrimination of movements into inversion at the ankle is associated with setting the degree of ankle inversion used during the swing phase of gait and increasing the risk of tripping (Waddington and Adams 2003). At the ankle, small differences in movement discrimination ability matter, because it is the large number of inversion movement repetitions (on average, 3.5 million steps per year) (Tudor-Locke et al. 2010) that potentially translates an increase in risk into a greater frequency of trip events.
The side-general nature of the effect of proprioceptive asymmetry observed here may reflect the “activity-specific nature of motor skills” (Provins 1997) or functional differences between preferred and non-preferred limbs during bilateral tasks (Goble et al. 2009). In daily functional activities, limbs on the preferred and non-preferred side of the body play different roles, particularly when tasks are executed in a “bilateral context” (Hart and Gabbard 1997). In task performance, the more distinct the stabilizing role of one limb, the stronger is the preference for the other limb (Grouios et al. 2009). Thus, for individuals with a strong unilateral preference, the non-preferred upper limb is usually used in statically stabilizing objects in a specific position for the preferred upper limb to manipulate in some way, for example removing a jar lid or hammering a nail. Similarly, the non-preferred lower limb is usually used in stabilizing the body for the preferred lower limb to swing, for instance kicking a ball or mounting a bicycle. It is hypothesized that, over time, conditioning of joints in the limbs with repetition could lead to use-dependent neuroplastic alterations of the hemisphere contralateral to each limb (Goble et al. 2009). Thus, joints in non-preferred limbs are more likely to receive more “positioning” practice, resulting in more accurate discrimination of movement, as observed here in strongly right-handed and right-footed participants. This finding is also consistent with Sainburg’s proposition that the non-preferred arm/hemisphere system is specialized for limb position control (Sainburg 2002, 2005), and the results of lower limb joints in the current study indicate that the non-preferred leg/hemisphere system follows the same rule.
Results obtained here may reflect limb asymmetry in sensory feedback processing (Goble and Brown 2008b). When performing voluntary movement, proprioception and vision are the two primary sources of sensory feedback (Goble and Brown 2008a). Several studies have shown that visual information has a different impact on movement control in the preferred/right versus non-preferred/left arm (e.g. Honda 1982; Roy and Elliott 1986), with greater importance of visual feedback in the movement control of the preferred/right arm (see Goble and Brown 2008a for review). This evidence suggests that preferred/right limb control tends to rely more on visual information than does the non-preferred/left side. Indeed, in daily activities, such as writing and kicking, preferred right limbs usually perform voluntary movements with vision available. Other evidence has indicated that although the left hemisphere plays a dominant role in movement control for right-handed individuals, the right hemisphere also has some specialized function in movement control (see Serrien et al. 2006 for review). For example, it has been proposed that the left hemisphere is specialized in the control of open-loop aspects of movement, based on well-developed motor programmes. In contrast, the right hemisphere is specialized in the control of the closed-loop aspects of movement, which are dependent on proprioceptive feedback (Haaland and Harrington 1989). Our results suggest that the right hemisphere may rely more on proprioceptive information for the control of non-preferred/left limbs, particularly when visual cues about target location are removed and only proprioceptive information is available.
However, the results of the present study are contrary to studies that did not show proprioceptive asymmetry between joints on the right and left sides of the body (Roy and MacKenzie 1978; Waddington and Adams 1999a; Bullock-Saxton et al. 2001; Naughton et al. 2002; Han et al. 2011). The lack of asymmetry in proprioceptive matching tasks has been largely attributed to differences in methodology, since active generation of target position in a criterion movement, for subsequent reproduction, has been thought to provide movement-related information that may enhance the matching performance of the preferred limb joint (Goble et al. 2006). In the current study, active movements were involved in both the familiarization sessions and testing, and results still showed a significant non-preferred side superiority in performance at each pair of joints, suggesting that other factors may underlie the lack of asymmetry observed in previous proprioceptive tests.
Firstly, most joint position reproduction (JPR) protocols for proprioception assessment usually use only 3–5 trials during the test (e.g. Bullock-Saxton et al. 2001; Zazulak et al. 2007; Adamo et al. 2009), and this may be insufficient to determine parameters in proprioceptive tests, as noted by Ashton-Miller (2000). A recent study estimated that 20 % of patients with proprioceptive deficits post-stroke would be missed if only 3 trials were used rather than 10 trials and that even when 10 trials were used, not all patients with proprioceptive deficits could be identified, suggesting that 10 is insufficient to set as a “gold standard” for a JPR protocol (Piriyaprasarth et al. 2009). Thus, the lack of limb asymmetry observed in JPR tests may be due to their low power arising from too few testing trials. In contrast, the AMEDA tests employed here used 50 trails at each site to assess the ability to discriminate between a set of different small movements. For analysis, the movement stimuli were considered as noise and signal in a pair-wise manner, to generate AUC values for the stimulus pairs 1–2, 2–3, 3–4 and 4–5. This nonparametric signal detection method (McNicol 2004) fulfils the requirement that a measure of the actual ability of proprioceptive mechanism to the sensorimotor system must take the effect of noise into account as a limit to proprioceptive ability (Clark et al. 1995). The greater sensitivity of the AMEDA tests may arise from a more stable mean AUC score and thus better reflect actual performance of proprioceptive mechanisms in the sensorimotor system. The high ICC scores of the reliability test at each joint support this notion.
Secondly, it has been argued that JPR tests of proprioception have low validity (Laszlo 1992; Ashton-Miller 2000), because the proprioceptive information available during target position generation and matching is not the same. The first difference between target position generation and matching is movement type. It has been suggested that in passive movement, since muscles are not active, fusimotor activity and the sensory feedback from muscle spindles are diminished. Thus, input from cutaneous receptors appears to play a greater role in sensory feedback. In contrast, in active movement control, fusimotor drive and muscle spindle feedback are both involved, although input from muscle spindles is considered to play a more dominant role (Gandevia et al. 1992; Zazulak et al. 2007). As a result, when a target joint position is passively generated for active matching, or vice versa, the brain may rely on different information from different receptors in the two phases, and the results may then reflect hemispheric specialization in the use of particular proprioceptive information at that joint. Further, information sources used in proprioception tests such as cutaneous receptors, joint receptors and muscle spindles differ in their relative contribution across different body sites (Proske and Gandevia 2009). Hence, when different proprioceptive testing protocols, such as passive–active ipsilateral or contralateral matching (see Goble 2010 for a review), are used to assess proprioceptive acuity at different joints, such as distal fingers/ankles and proximal shoulders/knees, both the source of information (Adamo and Martin 2009)—from ipsilateral or contralateral limb and the combination of proprioceptive sources—cutaneous receptors, joint receptors and muscle spindles, could possibly result in contradictory findings.
Another difference between target position generation used here and matching in JPR tests is that there is usually a physical stop at the end of the movement during target position generation, while the physical stop is removed during position matching. That is, during target position generation, information about movement extent and end position are both available, whereas only movement extent information is available when position matching. Although a movement extent/displacement matching strategy has been thought to be less effective than target position matching (see Goble et al. 2009 for detailed discussion), information about both limb movement extent and end position are needed for accurate judgement of limb movements (Magill and Parks 1983). In the present study, the proprioceptive information available during familiarization and testing is exactly the same, thus eliminating possible confounders between target position generation and matching in JPR tests. Therefore, the finding here of a side-general attribute in proprioceptive asymmetry suggests that the left limb/hemisphere system is integrating all normally available proprioceptive information from different receptors at different joints in active proprioceptive tasks.
Finally, failure to control for the degree of both handedness and footedness is another possible explanation for the lack of limb asymmetries in proprioceptive tasks reported previously. Studies using the same movement discrimination method did not find limb asymmetry (Waddington and Adams 1999a; Naughton et al. 2002; Han et al. 2011), presumably because side preference was not the main focus of the studies, and the degree of limb preference was not controlled. The present study controlled for the degree of both handedness and footedness and found that, for individuals with a strong right side preference, proprioceptive ability of the joints on the non-preferred/left side was consistently better than the joints on the preferred/right side. The handedness effect on sensorimotor performance has been investigated (Wang and Sainburg 2006; Goble et al. 2009; Adamo et al. 2012; Przybyla et al. 2012). For example, a recent study (Goble et al. 2009) recruited only strong left-handed individuals to perform upper limb position matching tasks and found that the sensorimotor performance of the left handers was the “mirror image” of their right-handed counterparts, implying right (non-preferred) arm/left hemisphere specialization in the use of proprioceptive feedback (Goble et al. 2009).
Another important finding of the present study is that significant positive and high correlations were found only between proprioceptive accuracy scores for the right and left sides at the same joint, whereas insignificant and low correlations were observed between scores at different joints. The finding that proprioceptive ability is site-specific in the current study extends previous lower limb movement discrimination results (Waddington and Adams 1999a) to sites in both upper and lower parts of the body. Thus, the results here do not support the “pre-existing global deficit” hypotheses (Goldie et al. 1994), rather the data are consistent with the hypothesis that sensitivity to the proprioceptive information that underlies movement control is site-specific.
In studies of patients with ankle injuries, Gauffin et al. (1988) and Waddington and Adams (1999b) have argued that the observation of poor performance of both injured and uninjured ankle favours a central motor programme view about bilateral joint movement control (Summers and Anson 2009). Similarly, bilateral deficits in fine motor control have been reported in patients with unilateral carpal tunnel syndrome (Fernández-de-las-Peñas et al. 2009). The correlations in the present study support this hypothesis, suggesting that there is likely to be a control programme that is common for the same joint on the two sides of the body and that the programme uses proprioceptive information from sources that are specific to those joints. This is true at the four pairs of joints tested in both the upper and lower limbs. The non-significant, low correlations between different body sites observed here suggest that the brain may utilize different programmes to integrate proprioceptive information from different sources at different joints. This notion is in line with previous findings that deficits of knee proprioception were not associated with poor ankle proprioception (Wang et al. 2008) and that ankle proprioceptive ability was improved by wobble board training, while this improvement did not alter knee proprioceptive ability (Waddington et al. 2000).
Overall, results here extend current understanding of the proprioceptive system by differentiating the side-general and site-specific effects of bilateral proprioceptive performance accuracy of multiple upper and lower limb joints. The results point to both a side-general effect and a site-specific effect in the integration of proprioceptive information during active movement tasks, whereby the non-preferred limb/hemisphere system is specialized in the utilization of the best proprioceptive sources available at each specific joint, but the relative contribution of the sources of proprioceptive information employed may differ at different sites of joints in the body.
References
Adamo D, Martin B (2009) Position sense asymmetry. Exp Brain Res 192:87–95
Adamo DE, Alexander NB, Brown SH (2009) The influence of age and physical activity on upper limb proprioceptive ability. Journal of Aging and Physical Activity 17:272–293
Adamo DE, Scotland S, Martin BJ (2012) Upper limb kinesthetic asymmetries: gender and handedness effects. Neurosci Lett 516:188–192
Ashton-Miller JA (2000) Proprioceptive thresholds at the ankle: Implications for the prevention of ligament injury. In: Lephart SM, Fu FH (eds) Proprioception and neuromuscular control in joint stability. Human Kinetics Publ, Champaign, pp 279–289
Bullock-Saxton JE, Wong WJ, Hogan N (2001) The influence of age on weight-bearing joint reposition sense of the knee. Exp Brain Res 136:400–406
Calguneri M, Bird HA, Wright V (1982) Changes in joint laxity occurring during pregnancy. Ann Rheum Dis 41:126–128
Clark FJ, Larwood KJ, Davis ME, Deffenbacher KA (1995) A metric for assessing acuity in positioning joints and limbs. Exp Brain Res 107:73–79
Cohen J (1992) A power primer. Psychol Bull 112:155–159
Fernández-de-las-Peñas C, Pérez-de-Heredia-Torres M, Martínez-Piédrola R, Llave-Rincón A, Cleland J (2009) Bilateral deficits in fine motor control and pinch grip force in patients with unilateral carpal tunnel syndrome. Exp Brain Res 194:29–37
Fleiss J (1986) The design and analysis of clinical experiments. Wiley, New York
Gandevia SC, McCloskey DI, Burke D (1992) Kinaesthetic signals and muscle contraction. Trends Neurosci 15:62–65
Gauffin H, Tropp H, Odenrick P (1988) Effect of ankle disk training on posture control in patients with functional instability of the ankle joint. Int J Sports Med 9:141–144
Gibson JJ (1979) The ecological approach to visual perception. Lawrence Erlbaum Associates, New Jersey
Goble DJ (2010) Proprioceptive acuity assessment via joint position matching: from basic science to general practice. Phys Ther 90:1176–1184
Goble DJ, Brown SH (2007) Task-dependent asymmetries in the utilization of proprioceptive feedback for goal-directed movement. Exp Brain Res 180:693–704
Goble DJ, Brown SH (2008a) The biological and behavioral basis of upper limb asymmetries in sensorimotor performance. Neurosci Biobehav Rev 32:598–610
Goble DJ, Brown SH (2008b) Upper limb asymmetries in the matching of proprioceptive versus visual targets. J Neurophysiol 99:3063–3074
Goble DJ, Brown SH (2009) Dynamic proprioceptive target matching behavior in the upper limb: effects of speed, task difficulty and arm/hemisphere asymmetries. Behav Brain Res 200:7–14
Goble DJ, Brown SH (2010) Upper limb asymmetries in the perception of proprioceptively determined dynamic position sense. J Exp Psychol-Hum Percept Perform 36:768–775
Goble DJ, Lewis CA, Brown SH (2006) Upper limb asymmetries in the utilization of proprioceptive feedback. Exp Brain Res 168:307–311
Goble DJ, Noble BC, Brown SH (2009) Proprioceptive target matching asymmetries in left-handed individuals. Exp Brain Res 197:403–408
Goldie PA, Evans OM, Bach TM (1994) Postural control following inversion injuries of the ankle. Arch Phys Med Rehabil 75:969–975
Grouios G, Hatzitaki V, Kollias N, Koidou I (2009) Investigating the stabilising and mobilising features of footedness. Laterality 14:362–380
Haaland KY, Harrington DL (1989) Hemispheric control of the initial and corrective components of aiming movements. Neuropsychologia 27:961–969
Han J, Waddington G, Anson J, Adams R (2011) A novel device for the measurement of functional finger pinch movement discrimination. Appl Mech Mater 66–68:620–625
Hart S, Gabbard C (1997) Examining the stabilising characteristics of footedness. Laterality 2:17–26
Honda H (1982) Rightward superiority of eye-movements in a bimanual aiming task. Q J Exp Psychol Sect Hum Exp Psychol 34:499–513
Jobbins B, Bird HA, Wright V (1979) A joint hyperextensometer for the quantification of joint laxity. Eng Med 8:103–104
Kalaycioglu C, Kara C, Atbasoglu C, Nalcaci E (2008) Aspects of foot preference: differential relationships of skilled and unskilled foot movements with motor asymmetry. Laterality 13:124–142
Kanegaonkar RG, Amin K, Clarke M (2012) The contribution of hearing to normal balance. J Laryngol Otol 126:984–988
Kuypers H (1982) A new look at the organization of the motor system. Prog Brain Res 57:381–403
Laszlo JI (1992) Motor control and learning: how far do the experimental tasks restrict our theoretical insight? In: Summers JJ (ed) Approaches to the study of motor control and learning, Elsevier, Amsterdam, pp 47–79
Lloyd AJ, Caldwell LS (1965) Accuracy of active and passive positioning of the leg on the basis of kinesthetic cues. J Comp Physiol Psychol 60:102–106
Magill RA, Parks PF (1983) The psychophysics of kinesthesis for positioning responses: the physical stimulus-psychological response relationship. Res Q Exerc Sport 54:346–351
McNicol D (2004) A primer of signal detection theory. Routledge, New York
Muaidi QI, Nicholson LL, Refshauge KM (2009) Do elite athletes exhibit enhanced proprioceptive acuity, range and strength of knee rotation compared with non-athletes? Scand J Med Sci Sports 19:103–112
Muller F, Kunesch E, Binkofski F, Freund H-J (1991) Residual sensorimotor functions in a patient after right-sided hemispherectomy. Neuropsychologia 29:125–145
Naughton J, Adams R, Maher C (2002) Discriminating overhead points of contact after arm raising. Percept Mot Skills 95:1187–1195
Naughton J, Adams R, Maher C (2005) Upper-body wobbleboard training effects on the post-dislocation shoulder. Physical Therapy in Sport 6:31–37
Nishizawa S (1991) Different pattern of hemisphere specialization between identical kinesthetic spatial and weight discrimination tasks. Neuropsychologia 29:305–312
Oldfield RC (1971) The assessment and analysis of handedness: the Edinburgh inventory. Neuropsychologia 9:97–113
Paillard J, Brouchon M (1968) Active and passive movements in the calibration of position sense. In: Freedman SJ (ed) The neuropsychology of spatially oriented behaviour. Dorsey Press, Homewood, pp 37–55
Piriyaprasarth P, Morris ME, Delany C, Winter A, Finch S (2009) Trials needed to assess knee proprioception following stroke. Physiother res int j res clinicians phys ther 14:6–16
Proske U, Gandevia SC (2009) The kinaesthetic senses. J Physiol-London 587:4139–4146
Provins KA (1997) The specificity of motor skill and manual asymmetry: a review of the evidence and its implications. J Mot Behav 29:183–192
Przybyla A, Good DC, Sainburg RL (2012) Dynamic dominance varies with handedness: reduced interlimb asymmetries in left-handers. Exp Brain Res 216:419–431
Roy EA, Elliott D (1986) Manual asymmetries in visually directed aiming. Can J Psychol-Revue Canadienne De Psychologie 40:109–121
Roy EA, MacKenzie CL (1978) Handedness effects in kinesthetic spatial location judgements. Cortex 14:250–258
Sainburg RL (2002) Evidence for a dynamic-dominance hypothesis of handedness. Exp Brain Res 142:241–258
Sainburg RL (2005) Handedness: differential specializations for control of trajectory and position. Exerc Sport Sci Rev 33:206–213
Serrien DJ, Ivry RB, Swinnen SP (2006) Dynamics of hemispheric specialization and integration in the context of motor control. Nat Rev Neurosci 7:160–167
Summers JJ, Anson JG (2009) Current status of the motor program: revisited. Hum Mov Sci 28:566–577
Swets JA (1996) Signal detection theory and ROC analysis in psychology and diagnostics: collected papers. Lawrence Erlbaum Associates Inc., Hillsdale
Swets JA, Tanner WP Jr, Birdsall TG (1961) Decision processes in perception. Psychol Rev 68:301–340
Swets JA, Dawes RM, Monahan J (2000) Better decisions through science. Sci Am 283:82–87
Symes M, Waddington G, Adams R (2010) Depth of ankle inversion and discrimination of foot positions. Percept Mot Skills 111:475–484
Tudor-Locke C, Johnson WD, Katzmarzyk PT (2010) Accelerometer-determined steps per day in US children and youth. Med Sci Sports Exerc 42:2244–2250
Vulcetic A, Holmes M, Adams R, Waddington G (2008) Bilateral associations of low-level unilateral performance: an unremarked aspect of limb control. J Mot Behav 40:479–483
Waddington G, Adams R (1999a) Ability to discriminate movements at the ankle and knee is joint specific. Percept Mot Skills 89:1037–1041
Waddington G, Adams R (1999b) Discrimination of active plantarflexion and inversion movements after ankle injury. Aust J Physiother 45:7–13
Waddington G, Adams R (2003) Football boot insoles and sensitivity to extent of ankle inversion movement. Br J Sports Med 37:170–174
Waddington G, Adams R, Jones A (1999) Wobble board (ankle disc) training effects on the discrimination of inversion movements. Aust J Physiother 45:95–101
Waddington G, Seward H, Wrigley T, Lacey N, Adams R (2000) Comparing wobble board and jump-landing training effects on knee and ankle movement discrimination. J Sci Med Sport 3:449–459
Wang JS, Sainburg RL (2006) Interlimb transfer of visuomotor rotations depends on handedness. Exp Brain Res 175:223–230
Wang L, Li JX, Xu DQ, Hong YL (2008) Proprioception of ankle and knee joints in obese boys and nonobese boys. Med Sci Monit: Int Med J Exp Clin Res 14:CR129–135
Wycherley AS, Helliwell PS, Bird HA (2005) A novel device for the measurement of proprioception in the hand. Rheumatology 44:638–641
Zazulak BT, Hewett TE, Reeves NP, Goldberg B, Cholewicki J (2007) The effects of core proprioception on knee injury—A prospective biomechanical-epidemiological study. Am J Sports Med 35:368–373
Acknowledgments
We would like to acknowledge the students and academic staff at the Shanghai University of Sport who gave their support and assistance in data collection for this study, and the University of Canberra for the provision of funds to conduct the research.
Conflict of interest
None.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 2.0 International License ( https://creativecommons.org/licenses/by/2.0 ), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Han, J., Anson, J., Waddington, G. et al. Proprioceptive performance of bilateral upper and lower limb joints: side-general and site-specific effects. Exp Brain Res 226, 313–323 (2013). https://doi.org/10.1007/s00221-013-3437-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00221-013-3437-0